Энергетический обмен головного мозга
4. АЭРОБНОЕ ОКИСЛЕНИЕ ГЛЮКОЗЫ В ГОЛОВНОМ МОЗГЕ И МЕХАНИЗМЫ ЕГО РЕГУЛЯЦИИ
В последние годы в литературе появились сведения о специфичности регуляторных механизмов, контролирующих метаболизм глюкозы в функционально различных органах и тканях в условиях интактного организма. Особое внимание исследователей сосредоточено на изучении соотношения активностей и механизмов контроля над теми фермента
ми, которые конкурируют за использование субстратов, стоящих в точках перекреста нескольких метаболических путей. Имеются лишь единичные публикации, посвященные исследованию механизмов, обеспечивающих высокую интенсивность энергетического обмена мозга in vivo, а также регуляции метаболизма мозга. Это объясняется сложной и гетерогенной структурой, наличием ГЭБ и рядом других причин, затрудняющих интерпретацию экспериментальных данных и экстраполяцию результатов, полученных in vitro, на мозг интактного животного. Многие аспекты этой важной и сложной проблемы требуют дальнейших углубленных нейрохимических исследований.
Пути утилизации глюкозы в мозге; гликолиз и механизмы, контролирующие его скорость
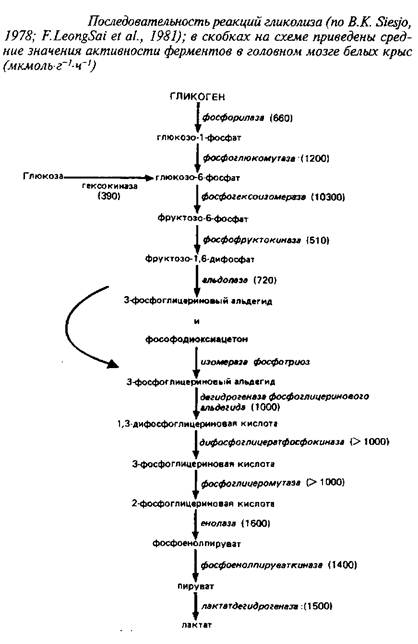
Последовательность реакций аэробного гликолиза, а также средние значения активностей ферментов, катализирующих отдельные стадии, приведены на схеме 1.
Из сопоставления активностей ферментов видно, что наиболее медленными реакциями, которые могут лимитировать скорость потока метаболитов по гликолитической цепи, являются гексокиназная и фосфофруктокиназная реакции.
Представление о количественном соотношении промежуточных компонентов гликолиза дают следующие средние результаты определения уровня метаболитов в головном мозге крыс:
|
Гликоген |
1,9 – |
3,8 |
|
УДФ-глюкоза |
0,08 – |
0,17 |
|
Глюкоза |
1,52 – |
3,70 |
|
Глюкозо-6-фосфат |
0,039- |
0,049 |
|
Фруктозо-б-фосфат |
0,017- |
0,023 |
|
Фруктозо – 1,6 – дифосфат |
0,010- |
0,017 |
|
Дигидроацетонфосфат |
0,024 | |
|
2-Глицероальдегидфосфат |
0,021- |
0,046 |
|
З-Фосфоглицерат |
0,100- |
0,085 |
|
2-Фосфоглицерат |
0,010- |
0,016 |
|
Фосфоенол п ируват |
0,035- |
0,097 |
|
Пируват |
0,120- |
0,190 |
|
Лактат |
1,26 – |
1,70 |
Гексокиназная реакция. Первым этапом на пути вовлечения свободной D-глюкозы, поступающей в мозг из крови, в разнообразные метаболические превращения служит реакция фосфорилирования, катализируемая гексокиназой. Для регуляции энергетического метаболизма в головном мозге гексокиназная реакция имеет особое значение, так как она является основным поставщиком глюкозо-6-фосфата, необходимого для дальнейших превращений. В других тканях, таких как печень, сердечная и скелетная мышцы и др., источником большей части глюкозо-6- фосфата служит реакция расщепления гликогена или реакции глюконеогенеза.
Гексокиназная реакция является доминирующим путем пополнения пула метаболитов гликолиза в мозге, поскольку, как уже упоминалось, глюкоза представляет собой основной энергетический субстрат в этой ткани. Окисление иных энергетических субстратов и ввод компонентов в гликолитическую цепь через другие реакции в нервной ткани не имеет существенного значения.
Все это позволяет рассматривать гексокиназную реакцию как первый пункт контроля над скоростью энергетического обмена в головном мозге. Лишь в экстремальных ситуациях – при резкой гипогликемии или в условиях чрезвычайно интенсивного гликолиза при кислородной недостаточности – лимитирующим этапом может стать транспорт глюкозы через ГЭБ.
Активность гексокиназьг мозга относительно невелика, особенно по сравнению с активностью других ферментов гликолиза; в среднем она составляет 350–450 мкмоль субстратаг-ч~. Эта величина в 5–10 раз превышает среднюю скорость поступления глюкозы в мозг. Более того, при сопоставлении активности фермента в различных тканях максимальные величины получены рядом авторов именно в экспериментах с головным мозгом.
Распределение фермента в нервных клетках неравномерно; с помощью гистохимических, иммунохимических методов, а также при исследовании обогащенных препаратов установлена более высокая активность гексокиназы в нейронах, особенно в си-наптических окончаниях, по сравнению с олигодендроглией.
Интересная особенность отмечена при изучении распределения фермента между компартментами клетки: в отличие от ряда других тканей в мозге основная часть гексокиназы сосредоточена не в цитоплазме, а в митохондриях. В связывании фермента с внешней митохондриальной мембраной участвует специфический белок, детальные исследования свойств которого указывают на идентичность его с белком, формирующим поры. На прочность взаимодействия гексокиназы с мембранным белком оказывает влияние фосфолипидный компонент мембраны. Причины такого своеобразного внутриклеточного распределения гексокиназы в мозге пока не совсем ясны, но имеются предположения, что такая локализация обеспечивает более быстрое и эффективное фосфорилирование глюкозы за счет АТФ, синтезированного в митохондриях.
Необходимо отметить, что из четырех известных изофеоментов гексокиназы – ГКГ, ГКП, ГКШ, ГК1У в мозге встречается лишь первые два, причем на долю ГКТ приходится около 90% суммарной активности. Именно для этих двух изоферментов наиболее выражена способность связываться с внешней митохондриальной мембраной.
Связывание гексокиназы – обратимый процесс; на соотношение между связанной и солюбилизированной формами фермента влияет рад факторов, в первую очередь – отношение АТФ/АДФ, концентрация неорганического фосфата и глкжо-зо-6-фосфата, а также уровень свободных: жирных кислот. Действие указанных факторов проявляется в пределах их физиологических концентраций; это дало Вильсону основание предположить, что изменение субклеточного распределения гексокиназы служит важным механизмом регуляции активности фермента in vivo.
Другие рефераты на тему «Медицина»:
Поиск рефератов
Последние рефераты раздела
- Особенности лечения и тракционно-экстензионной терапии на аппарате Kinetrac KNX-7000
- Остеохондроз, методики лечения
- Тракционно-экстензионная терапия у больных остеохондрозом пояснично-крестцового отдела позвоночника
- Болезни, возникающие от курения. Профилактика курения
- Болезни органов дыхания
- Болезни желчевыводящих путей и печени
- Анатомия и физиология артерий нижних конечностей. Этиология и патогенез