Денервация и регенерация синаптических связей
Рецепторы АХ, появляющиеся во внесинаптических областях, не просто диффундируют от области исходной концевой пластинки. Этот факт впервые был установлен в экспериментах Катца и Миледи, в которых мышца лягушки была разрезана на две части. Содержащие ядра фрагменты мышцы, физически отделенные от исходной концевой пластинки, не только выживали, но и развивали повышенную чувствительность к АХ. Глав
ный вывод, который следует из этих экспериментов, заключается в том, что новые АХ рецепторы способны синтезироваться в экстрасинаптических участках денервированных мышц.
Синтез и деградация рецепторов в денервированной мышце
Ценным методом для изучения распределения и реорганизации АХ рецепторов является их мечение радиоактивным бунгаротоксином, который прочно и с высокой специфичностью связывается с АХ рецепторами. Мечение нормальной и денервированной мыши бунгаротоксином подтвердило, что число и распределение центров связывания токсина изменяется после денервации. В постсинаптической мембране нормальной мышцы определяется около 104 сайтов связывания токсина на мкм2, в сравнении с менее чем 10/мкм2 во внесинаптической области. После денервации число АХ рецепторов во внесинаптических участках возрастает до 103/мкм2, тогда как в синаптической зоне плотность центров связывания бунгаротоксина меняется незначительно.
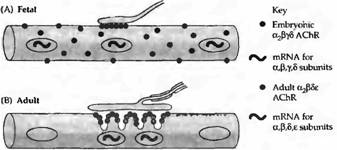
Рост числа АХ рецепторов в денервированной мышце может быть обусловлен их усиленным синтезом. Действительно, скорость появления новых рецепторов заметно повышается после денервации, и вещества, блокирующие синтез белка (такие как актиномицин или пуромииин), предупреждают этот эффект. Нозерн блоттинг и гибридизация in situ показывают, что в нормальной мышце мРНК для субъединиц АХ рецептора синтезируют только несколько ядер, локализующихся в непосредственной близи от концевой пластинки. В противоположность этому, при денервации гены АХ рецептора активируются по всей протяженности мышечного волокна (рис. 4).
Денервация оказывает также влияние на композицию субъединиц и скорость деградации АХ рецепторов. Во взрослой мышце синаптические и экстрасинаптические АХ рецепторы содержат субъединицу с периодом полураспада около 10 дней. После денервации период полураспада рецепторов, содержащих субъединицу и сохраняющихся в области концевой пластинки, уменьшается до 3 дней. Оборот рецепторов может быть вновь замедлен реиннервацией или повышением концентрации внутриклеточного цАМФ и последующей активацией протеинкиназы А.
Новые рецепторы, синтезированные в денервированной мышие (синаптические или внесинаптические), похожи на эмбриональные. Они содержат субъединицу и имеют оборот с периодом полураспада 1 день. Эта скорость оборота АХ рецепторов может быть снижена экзогенным АТФ, действующим через пуринергические рецепторы.
Роль инактивации мышцы в денервационной гиперчувствительности
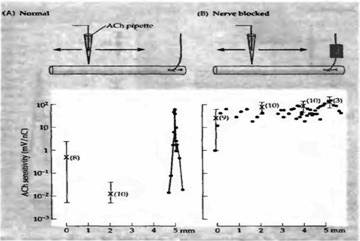
Каким образом перерезка двигательного нерва приводит к появлению новых АХ рецепторов — через инактивацию мышцы или через какие-то другие механизмы? Для исследования этой проблемы Ломо и Розенталь блокировали проведение импульсов по двигательному нерву крысы аппликацией местного анестетика или дифтерийного токсина. Вещества апплицировались с помощью манжеты, наложенной на участок нерва на некотором расстоянии от мышцы. Этот метод приводил к эффективной инактивации мышцы, так как двигательные импульсы не проводились дальше манжеты. Тестовая стимуляция нерва дистальнее блокированного участка приводила к обычному сокращению мышцы. Кроме того, происходила нормальная генерация миниатюрных потенциалов концевой пластинки, свидетельствуя, что синаптическая передача была интактна. После 7 дней блока нерва мышца становилась гиперчувствительной (рис. 5). Другими экспериментами было показано, что новые внесинаптические рецепторы появляются тогда, когда нервно-мышечная передача блокирована долговременной аппликацией кураре или бунгаротоксина. Эти результаты показали, что денервационная гиперчувствительность связана с потерей синаптической активации мышцы.
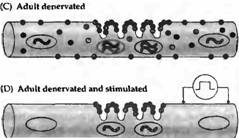
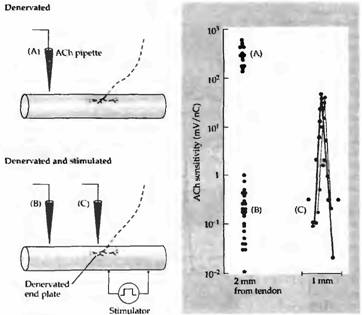
Важность мышечной активности как фактора, контролирующего реакцию мышцы на АХ, была подтверждена в экспериментах, в которых гиперчувствительные денервированные мышцы крысы прямо стимулировали через имплантированные электроды. Повторяющаяся в течение нескольких дней стимуляция мышц сокращала область, чувствительную к АХ, до уровня исходного синаптического участка (рис. 4D и 6). Спонтанная сократительная активность мышцы в этих условиях была слишком низкой для того, чтобы обратить эффекты денервации на распределение АХ рецепторов
|
Рис. 4. Синтез и распределение АХ рецепторов в мышце крысы. (А) В эмбриональных мышцах АХ рецептора экспрессируется в ядрах по всей протяженности мышечного волокна. Эмбриональная форма рецептора определяется по всей поверхности мышечного волокна и аккумулируется в зоне иннервации. (В) Во взрослых мышцах мРНК экспрессируется только в ядрах, располагающихся непосредственно под концевой пластинкой. . (С) В денервированных взрослых мышцах ядра непосредственно под концевой пластинкой экспрессируют; все другие ядра экспрессируют эмбриональную. Эмбриональные АХ рецепторы найдены на всей поверхности мышечных волокон (обеспечивая таким образом денервационную гиперчувствительность), включая постсинаптическую мембрану; взрослая форма рецептора ограничена участком концевой пластинки. (D) При прямой стимуляции денервированных мышц |
паттерн экспрессии АХ рецептора напоминает таковой в иннервированном мышечном волокне. |
|
Рис. 5. В мышце крысы блокирование нервной проводимости приводит к появлению новых АХ рецепторов. (А) В нормальной мышце чувствительность к АХ ограничена участком концевой пластинки. (В) После блокады двигательного нерва в течение 7 дней местным анестетиком чувствительность к АХ распространяется по всей поверхности мышечного волокна. |
| |||
|
Рис. 6. Устранение гиперчувствительности в денервированной мышце крысы прямой стимуляцией мышечных волокон. (А) Увеличение чувствительности во внесинаптической части мышечного волокна после 14 дней денервации. (В) Чувствительность внесинаптического участка денервированной мышцы, сохранявшейся в течение 7 дней без стимуляции, а затем стимулированной с перерывами в последующие 7 дней. Эта процедура устраняла денервационную гиперчувствительность. (С) Чувствительность к АХ двух стимулированных волокон той же мышцы рядом с денервированными участками концевой пластинки. В стимулированной мышце высокая чувствительность к АХ ограничена этим участком. |
| |||
Другие рефераты на тему «Медицина»:
Поиск рефератов
Последние рефераты раздела
- Особенности лечения и тракционно-экстензионной терапии на аппарате Kinetrac KNX-7000
- Остеохондроз, методики лечения
- Тракционно-экстензионная терапия у больных остеохондрозом пояснично-крестцового отдела позвоночника
- Болезни, возникающие от курения. Профилактика курения
- Болезни органов дыхания
- Болезни желчевыводящих путей и печени
- Анатомия и физиология артерий нижних конечностей. Этиология и патогенез