Денервация и регенерация синаптических связей
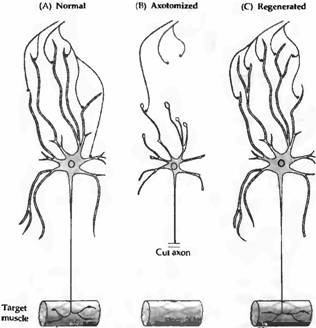
Через несколько часов после перерезки начинается рост новых аксональных отростков, происходящих от перерезанного конца нервного волокна, и процесс регенерации. Если нейрон успешно восстановил контакт с мишенью, клеточное тело приобретает исходный внешний вид. Хроматолиз происходит не только в периферических нервах, но также и после повреждения аксонов в центральной нервной системе.
В орга
низме взрослых животных реакция нейронов, не восстановивших контактов со своими мишенями, может быть различной. Пересечение аксонов оптического нерва приводит к быстрой гибели ганглиозных клеток сетчатки. Большинство клеток переднего базального таламуса выживают после повреждений коры, хотя при этом они существенно атрофируются. Чувствительные нейроны задних корешков спинного мозга и мотонейроны после перерезки их аксонов в периферическом нерве, как правило, погибают. Немногие выжившие при этом клетки в значительной степени атрофируются. Большинство аксотомированных клеток автономного ганглия выживают, но они уменьшаются в размерах и становятся менее чувствительными к ацетилхолину.
Ретроградные транссинаптические эффекты аксотомии
Аксотомия может также вызывать изменения в нейронах, посылающих синаптический сигнал к поврежденной клетке. Например, после аксотомии клеток автономного ганглия цыпленка, крысы или морской свинки, синаптические сигналы в клетках ганглия становятся менее эффективными. Это отчасти является результатом сниженной чувствительности аксотомированной клетки к нейротрансмиттеру ацетилхолину. Кроме того, как результат ретроградных транссинаптических влияний, происходит ретракция пресинаптических окончаний, а оставшиеся терминали освобождают меньше квантов трансмиттера (рис. 2). Таким образом, повреждение нейрона нарушает его способность поддерживать высокоэффективный пресинаптический сигнал. Ротшенкер показал и другой пример ретроградного транссинаптического влияния, наблюдаемый в мотонейронах лягушки и мыши. После перерезки моторного нерва на одной стороне тела происходит формирование новых синапсов и спраутинг аксонов интактных мотонейронов, иннервируюших соответствующую мышцу другой стороны тела. Сигнал распространяется от аксотомированных нейронов, пересекает спинной мозг и воздействует на неповрежденные мотонейроны на другой стороне животного. Важно отметить, что мотонейроны, иннервирующие другие мышцы, при этом не затрагиваются.
Трофические субстанции и эффекты аксотомии
Ряд эффектов аксотомии — хроматолиз, атрофия нейронов и гибель клеток — является результатом утраты трофических веществ, продуцируемых тканями-мишенями и транспортируемых по аксону ретроградно с периферии по направлению к телу клетки. Хорошим примером этого является действие фактора роста нервов (NGF) на сенсорные и симпатические нейроны. Так, в автономном ганглии морской свинки эффекты аксотомии воспроизводятся подкожными инъекциями антител к фактору роста нервов или блокированием ретроградного транспорта в постганглионарных нервах. Напротив, эффекты аксотомии в значительной мере предотвращаются аппликацией NGF на ганглий. Подобным же образом ретроградная транссинаптическая атрофия может быть результатом снижения продукции трофических факторов поврежденными нейронами.
|
Рис. 2. Атрофия клеток аксотомированного автономного ганглия и потеря пресинаптических входов. (А) Нормальный нейрон. (В) Через несколько дней после аксотомии нейроны атрофируются большая часть дендритов приобретает варикозные расширения. Многие пресинаптические окончания подвергаются ретракции, а оставшиеся освобождают меньше трансмиттера. (С) Если постганглионарный аксон регенерирует и реиннервирует свою периферическую мишень, свойства клетки и синаптических входов восстанавливаются. |
|
Эффекты денервации на постсинаптические клетки
Нервно-мышечный синапс является удобной моделью для изучения механизмов синаптической передачи, которые приложимы не только к периферическим, но и к центральным синапсам. Сходным образом процессы, происходящие в денервированных мышцах, могут быть хорошим аналогом того, к чему приводит устранение синаптических входов в нейронах ЦНС.
Денервированная мышечная мембрана
В конце девятнадцатого столетия в денервированной скелетной мышце был обнаружен феномен спонтанных асинхронных сокращений, названных фибрилляциями. Фибрилляции инициировались самой мышечной мембраной, а не АХ, хотя большинство спонтанных потенциалов действия, вызывающих фибрилляцию, исходило из участка бывшей концевой пластинки. Начало фибрилляций у крыс, морских свинок или кроликов наблюдалось через 2-5 дней после денервации, тогда как у обезьян и человека этот латентный период занимал более недели.
К моменту начала фибрилляций волокна мышц млекопитающих становятся гиперчувствительными к различным химическим агентам. Это значит, что концентрация веществ, требуемых для возбуждения мышцы, снижается в сотни или тысячи раз. Например, денервированная скелетная мышца млекопитающего примерно в 1000 раз более чувствительна к АХ, добавленному непосредственно в омывающий раствор или инъецированному в артерию, снабжающую мышцу, чем нормально иннервированная мышца17). Потенциал действия в денервированных мышцах также меняется, становясь более устойчивым к тетродотоксину, яду рыбы футу, который блокирует натриевые каналы . Это изменение обусловлено повторным появлением тетродотоксин-резистентных натриевых каналов, которые являются превалирующей формой каналов в незрелой мышце. В денервированной мышце происходят и другие изменения, такие как постепенная атрофия или опустошение мышечных волокон19
|
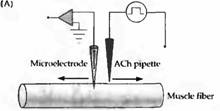
Рис. 3. После денервации в мышце кошки появляются новые АХ рецепторы. (А) Порции АХ наносятся из пипетки, заполненной АХ, на разные участки поверхности мышечного волокна, изменения мембранного потенциала при зтом регистрируются внутриклеточным микроэлектродом. (В) В мышечном волокне с интактной иннервацией АХ вызывает ответ только вблизи концевой пластинки. (С) После 14 дней денервации мышечное волокно отвечает на АХ по всей своей длине. |
|
Появление новых АХ рецепторов после денервации или длительной инактивации мышцы
Гиперчувствительность к ацетилхолину объясняется повышением числа и изменением распределения АХ рецепторов в денервированной мышце. Этот факт был установлен в результате экспериментов, в которых записывали изменения мембранного потенциала мышечного волокна во время ионофоретической аппликации АХ из микропипетки на небольшие участки мышечной мембраны. В нормально иннервированной мышце лягушки, змеи или млекопитающего чувствительным к АХ был только участок концевой пластинки — там, где нервное волокно образует синапс. Остальная мышечная мембрана имела очень низкую чувствительность к медиатору. После денервации область, реагирующая на АХ, значительно увеличивалась, достигая практически равномерной чувствительности поверхности мышцы к АХ (рис. 3). У млекопитающих это занимало около недели; в мышце лягушки изменения были выражены меньше и развивались заметно дольше.
Другие рефераты на тему «Медицина»:
Поиск рефератов
Последние рефераты раздела
- Особенности лечения и тракционно-экстензионной терапии на аппарате Kinetrac KNX-7000
- Остеохондроз, методики лечения
- Тракционно-экстензионная терапия у больных остеохондрозом пояснично-крестцового отдела позвоночника
- Болезни, возникающие от курения. Профилактика курения
- Болезни органов дыхания
- Болезни желчевыводящих путей и печени
- Анатомия и физиология артерий нижних конечностей. Этиология и патогенез