Масс-спектрометрическая оценка уровня включения дейтерия и углерода-13 в молекулы аминокислот бактериальных объектов
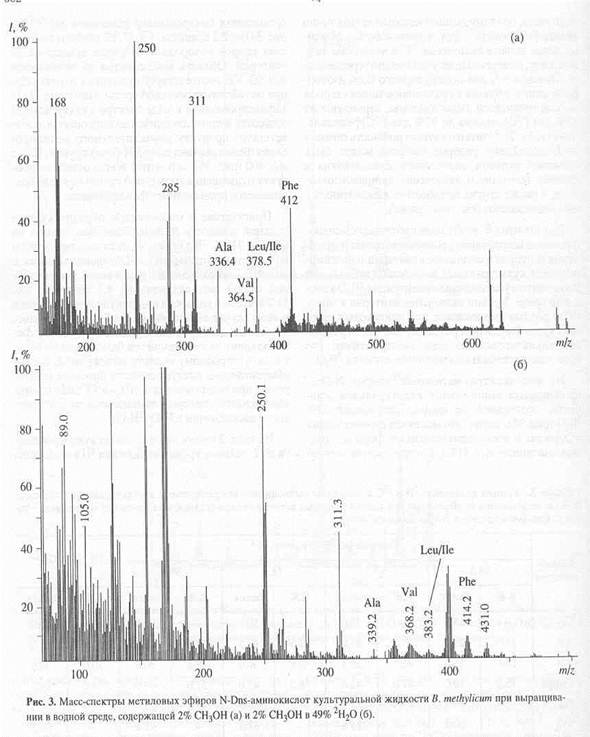
Присутствие в масс-спектре образца культуральной жидкости B. methylicum, полученной на среде с 73.5% 2Н2О (рис. 4) пика молекулярного иона метилового эфира N-Dns-фенилаланина с m/z 416.1 указывает на увеличение молекулярной массы фенилаланина на 4.1 единицу, т. е., 51.2% атомов водорода в молекуле фенилала
нина в этом случае замещены на дейтерий. Очевидно, что вышеобозначенные атомы дейтерия включились в молекулу фенилаланина за счет процесса биосинтеза de novo, т. е. по углеродному скелету молекулы. К легко обмениваемым следует отнести протоны (дейтероны) при гетероатомах в NH2- и СООН- группах аминокислот, которые замещаются за счёт лёгкости диссоциации в Н2О (2Н2О).
Таблица 2
Уровни включения 2Н и 13С в молекулы аминокислот, секретируемых в культуральную жидкость (КЖ) B. methylicum и M. flagellatum, и в аминокислотные остатки белков (данные получены для метиловых эфиров N-Dns-аминокислот и N-Cbz-аминокислот)
|
Аминокислоты | Содержание 2Н2О в среде, %* 24.5 49.0 73.5 98.0 КЖ белок КЖ белок КЖ белок КЖ белок |
1%13СН3ОН** КЖ белок | ||||
|
Глицин |
- 15.0 |
- 35.0 |
- 50.0 |
- 90.0 |
60.0 90.0 | |
|
Аланин |
24.0 20.0 |
37.5 45.0 |
62.5 62.5 |
77.5 97.5 |
35.0 95.0 | |
|
Валин |
20.0 15.0 |
46.3 36.3 |
43.8 50.0 |
58.8 50.0 |
50.0 50.0 | |
|
Лейцин/изолейцин |
15.0 10.0 |
47.0 42.0 |
46.0 45.0 |
51.0 49.0 |
38.0 49.0 | |
|
Фенилаланин |
15.0 24.5 |
27.5 37.5 |
51.2 50.0 |
75.0 95.0 |
95.0 80.5 | |
|
Тирозин |
- 20.0 |
- 25.6 |
- 68.8 |
- 92.8 |
- 53.5 | |
|
Серин |
- 15.0 |
- 36.7 |
- 47.6 |
- 86.6 |
- 73.3 | |
|
Аспарагиновая кислота |
- 20.0 |
- 36.7 |
- 60.0 |
- 66.6 |
- 33.3 | |
|
Глутаминовая кислота |
- 20.0 |
- 40.0 |
- 53.4 |
- 70,0 |
- 40.0 | |
|
|
- 10.0 |
- 35.3 |
- 40.0 |
- 58.9 |
- 54.4 | |
Из табл. 2 видно, что условиях ауксотрофности по L-лейцину уровни включения 2Н в молекулы лейцина/изолейцина ниже, чем для фенилаланина. Отмеченная особенность отчётливее всего проявляется на среде с максимальной концентрацией 2Н2О. Ещё раз этот результат подтвердили рис. 5, где показан масс-спектр [2Н]аминокислот культуральной жидкости в виде метиловых эфиров N-Dns-производных аминокислот после выращивания бактерий B. methylicum в указанных условиях. Видно, что величина m/z 418.0 пика молекулярного иона метилового эфира N-Dns-фенилаланиа увеличивается по сравнению с контрольными условиями на 6 единиц, что соответствует замещению 75% от общего количества атомов водорода в молекуле. В отличие от фенилаланина уровень включения дейтерия в лейцин/изолейцин составил 51.0%, а в валин - 58.8%. Примечательно, что в масс-спектре этого образца фиксируется пик обогащённого дейтерием бензильного фрагмента при m/z 97.0 (вместо m/z при 91.0 в контроле), что указывает на частичную локализацию атомов дейтерия в молекуле фенилаланина в положениях С2-С6 ароматического кольца и сопредельном с ними положении при углеродном атоме b. Несмотря на то, что в остальных опытах пики бензильных фрагментов не были зафиксированы, логично предположить, что при других концентрациях 2Н2О дейтерий также включается в ароматическое кольцо фенилаланина, так как в 2Н2О метаболизм у штамма B. methylicum не претерпевает существенных изменений [25].
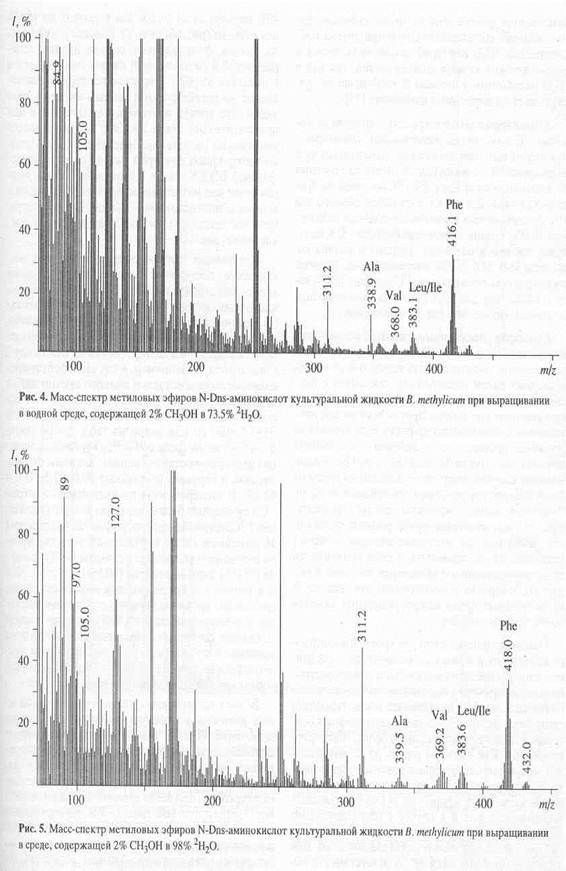
Аналогичная закономерность в уровнях включения 13С в молекулы аминокислот, связанных с ауксотрофным метаболизмом, проявляется при выращивании L-изолейцинзависимого штамма M. flagellatum на среде с 1% (13С)метанолом. Как видно из табл. 2, в отличие от наблюдаемого для [13С]фенилаланина (уровень изотопного включения - 95.0%), уровни включения изотопа 13С в молекулы лейцина/изолейцина, аланина и валина составили 38.0; 35.0; 50.0% соответственно. Уровень изотопного включения для [13C]глицина (60%) хотя и выше, чем для трёх последних аминокислот, но намного ниже, чем для фенилаланина.
Суммируя полученные данные по уровням включения 2Н-и 13С в молекулы секретируемых аминокислот, можно сделать вывод о сохранении минорных путей метаболизма, связанных с биосинтезом лейцина и метаболически родственных с ним аминокислот de novo. Другим логическим объяснением наблюдаемого эффекта, если принять во внимание происхождение лейцина и изолейцина по различным путям биосинтеза, может быть ассимиляция клеткой немеченого лейцина из среды на фоне биосинтеза меченого изолейцина de novo. Учитывая данные эффекты следует подчеркнуть, что использование ауксотрофных форм микроорганизмов для получения изотопномеченых аминокислот не оправдывает себя практически из-за множественного включения изотопов в молекулы. Напротив, использование для этих целей прототрофных форм микроорганизмов кажется более перспективным.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики