Масс-спектрометрическая оценка уровня включения дейтерия и углерода-13 в молекулы аминокислот бактериальных объектов
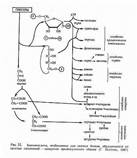
Общие принципы изучения уровней изотопного включения в молекулы аминокислот при данном способе введения метки были продемонстрированы на примере анализа сложных мультикомпонентных смесей, полученных после гидролиза суммарных белков биомассы метилотрофных бактерий, а также индивидуального белка – бактериородопс
ина, выполняющего роль АТФ-зависимой транслоказы в клетках галофильной бактерии Halobacterium halobium. Как видно из рис. 6, до десяти аминокислот могут быть идентифицированы в гидролизате белка B. methylicum по пикам молекулярных ионов метиловых эфиров их N-Dns-производных аминокислот.
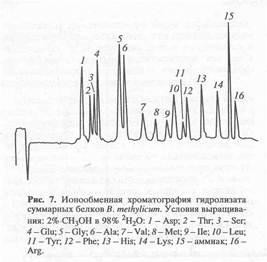
Как и в случае с секретируемыми аминокислотами, пики М+. соответствовали смесям изотопнозамещённых форм аминокислот. Для лизина и тирозина пики М+. соответствовали метиловым эфирам ди-производных аминокислот - a, e-ди-Dns-лизину (с М+. при m/z 631.0) и О, N-ди-Dns-тирозину (с М+. при m/z 663.9). Уровни изотопного включения дейтерия в молекулы аминокислот при содержании 2Н2O в ростовой среде 49% варьируют от 25.6% для тирозина до 45.0% для аланина (рис. 6 б и табл. 2). В молекулах глицина, валина, фенилаланина, серина, лизина, аспарагиновой и глутаминовой кислот они находятся в пределах 35 - 40%. Что касается других аминокислот, не детектируемых данным методом, очевидно, что уровни изотопного включения в них приблизительно такие же. Это подтверждается данными по разделению белковых гидролизатов метилотрофных бактерий методами обращённо-фазовой ВЭЖХ в виде N-Cbz-производных аминокислот или метиловых эфиров их N-Dns-производных аминокислот и ионнообменной хроматографии, где детектируется уже 15 аминокислот (см., например, рис. 7).
Полученные данные свидетельствуют о возможности достижения максимальных уровней включения стабильных изотопов 2Н и 13С в аминокислотные остатки суммарных белков биомассы (за исключением лейцина/изолейцина и валина, сниженные уровни включения для которых объясняются эффектом ауксотрофности по L-лейцину и по L-изолейцину). Например, в случае с дейтерированными аминокислотами полного замещения на стабильные изотопы удалось достичь за счет использования в качестве источника дейтерия 98% 2Н2О (табл. 2). Как видно из табл. 2, при росте B. methylicum на среде с 98% 2Н2О, уровни включения дейтерия в остатки глицина, аланина, фенилаланина и тирозина составляют 90.0; 97.5; 95.0 и 92.8%. В экспериментах по включению изотопа 13С в суммарные белки биомассы за счёт утилизации (13С)метанола метилотрофными бактериями M. flagellatum также наблюдались высокие уровни изотопного включения в глицине (90%), аланине (95.0%) и фенилаланине (80.5%) (табл. 2). Как и в случае с секретируемыми аминокислотами, сниженные уровни включения стабильных изотопов в лейцине/изолейцине (49%), а также в метаболически связанных с ним аминокислотах в этих условиях могут быть объяснены эффектом ауксотрофности штамма по L-изолейцину, который добавляли в ростовую среду в немеченом виде.
Во всех экспериментах по включению стабильных изотопов в молекулы аминокислот уровни включения 2Н и 13С в метаболически связанные аминокислоты обнаружили определённую коррелляцию. Так, уровни изотопного включения для валина и лейцина (семейство пирувата), фенилаланина и тирозина (семейство ароматических аминокислот) коррелируют (см. табл. 2). Уровни изотопного включения для глицина и серина (семейство серина), аспарагиновой кислоты и в лизина (семейство аспарагина) также имеют близкие величины. Из данных табл. 2 видно, что уровни изотопного включения секретируемых аминокислот и соответствующих аминокислотных остатков суммарного белка при выращивании бактерий на средах с одинаковым изотопным насыщением, в целом, также коррелируют. Причина некоторых наблюдаемых расхождений в уровнях включения изотопов в молекулы аминокислот до конца не изучена.
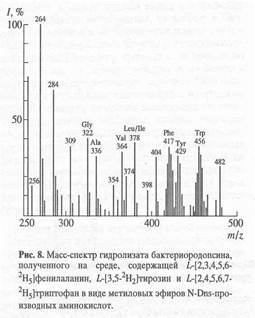
Данный биосинтетический подход показал хорошие результаты по изучению введения дейтериевой метки в молекулу бактериородопсина, выращенного на среде, содержащей L-[2,3,4,5,6-2Н]фенилаланин, L-[3,5-2Н]тирозин и L-[2,4,5,6,7-2Н]триптофан (рис. 8). Как видно из рис. 8, в масс-спектре дериватизованного гидролизата бактериородопсина детектируются пики, соответствующие молекулярным ионам обогащённых дейтерием метиловых эфиров N-Dns-фенилаланина с молекулярным ионом при m/z 417 (ср. m/z 412 для немеченого производного фенилаланина), N-Dns-тирозина с М+. при m/z 429 (ср. m/z 428 для производного тирозина) и N-Dns-триптофана с М+. при m/z 456 (ср. m/z 451 для производного триптофана). Все они отвечают смеси изотнопозамещённых форм аминокислот, различающихся количеством атомов водорода, замещённых на дейтерий. Множественный характер включения дейтерия свидетельствует о возможном вкладе биосинтеза de novo в уровни дейтерированности ароматических аминокислот, но также не исключено, что он определяется самим способом получения изотопномеченых молекул. Кроме вышеобозначенных аминокислот в масс-спектре фиксируются пики молекулярных ионов метиловых эфиров -N-Dns-глицина (m/z 322), N-Dns-аланина (m/z 336), N-Dns-валина (m/z 364) и N-Dns-лейцина/изолейцина (m/z 378). Как и следовало ожидать, эти аминокислотные остатки в бактериородопсине не содержат дейтерия.
Таким образом, проведённые исследования продемонстрировали эффективность масс-спектрометрии электронного удара N-Cbz-производных аминокислот и метиловых эфиров N-Dns-производных аминокислот для исследования уровней изотопного обогащения молекул аминокислот в составе их мультикомпонентных смесей, полученных биосинтетически с использованием микроорганизмов. Метод незаменим для изучения состава пула аминокислот, секретируемых в культуральные жидкости штаммов-продуцентов, выращенных на средах со стабильными изотопами.
Экспериментальная часть
В работе использовали D, L-аминокислоты (Reanal, Венгрия), аденозин- и уридин-5-монофосфаты (Sigma, США), панкреотическую телячью дезоксирибонуклеазу I (Fluka Chemie AG, Швейцария), додецилсульфат натрия (Chemapol, Чехо-Словакия). L-[2,3,4,5,6-2Н5]фенилаланин (90 ат.% 2Н), L-[3,5-2Н2]тирозин (96 ат.% 2Н) и L-[2,4,5,6,7-2Н5]триптофан (98 ат.% 2Н) (способы получения указаны в работах [34, 35]), были предоставлены А. Б. Пшеничниковой (МИТХТ им. М. В. Ломоносова). Для синтеза производных аминокислот использовали N-диметиламинонафталин-5-сульфохлорид (дансилхлорид) (Sigma, США), бензилоксикарбонилхлорид (Войковский химзавод, РФ) и диазометан, получаемый из N-нитрозометилмочевины (Merck, Германия).
Исследования проводили с генетически маркированными штаммами бактерий, полученными из коллекции культур Всероссийской коллекции промышленных микроорганизмов (ВКПМ) Государственного научно-исследовательского института генетики и селекции промышленных микроорганизмов:
Brevibacterium methylicum ВКПМ В 5652, L-лейцинзависимый штамм факультативных метилотрофных бактерий, продуцент L-фенилаланина;
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики