Как возникает нервный импульс
Попробуйте сами, не читая дальше, посмотреть эти данные и придумать механизм возникновения ПД, для чего фактически достаточно заполнить многоточия в помещенной ниже схеме.

Сделав это, вы получите, по существу, формулировку так называемой натриевой гипотезы потенциала действия, авторами которой является уже знакомый нам
Ходжкин и его сотрудник Хаксли.
Суть этой гипотезы заключается в том, что при возбуждении мембрана не просто полностью утрачивает избирательную проницаемость, а меняет ее: из проницаемой главным образом для К+ мембрана становится проницаемой главным образом для Ка+. А так как натрия снаружи больше, то он «лезет» внутрь и перезаряжает мембрану – получается как бы «ПП наоборот». Этим легко объясняется овершут.
Как видите, натриевая гипотеза настолько органично вписывается в мембранную теорию, что может показаться странным, что в противоречии существования овершута с мембранной теорией не могли разобраться свыше десяти лет. Но этот факт легко объясняется, если вспомнить, что овершут был обнаружен в 1939 г. Как когда-то первая мировая война помешала Бернштейну исследовать гипотезу «дырок», так начавшаяся 1 сентября 1939 г., вторая мировая война заставила Ходжкина и Хаксли отложить свои исследования. Во время войны Ходжкин работал в министерстве ВВС и занимался разработкой радиолокационных станций. Лишь после войны, в конце 40-х – начале 50-х годов, натриевая гипотеза превращается в стройную, экспериментально доказанную и логически безупречную натриевую теорию.
От гипотезы до теории
Первая экспериментальная проверка гипотезы была предпринята А. Ходжкином и Б. Катцем в 1949 г. Исследователи решили посмотреть, как влияет на величину ПД изменение концентраций калия и натрия в наружной среде. Они показали, что амплитуда ПД зависит от изменений концентрации натрия так же, как величина ПП зависит от концентрации калия, т.е. подчиняется формуле Нернста. Это было хотя и косвенное, но сильное подтверждение натриевой гипотезы: ведь это и означает, что в возбужденной мембране калиевая проницаемость сменяется натриевой. Используя более тонкий метод для сопоставления проницаемости мембраны для калия и натрия, Ходжкин и Катц установили, что в покое мембрана примерно в 25 раз более проницаема для кaлия, чем для натрия, а при возбуждении, наоборот, проницаемость для натрия становится примерно в 20 раз выше, чем для калия.
Основная идея натриевой гипотезы была подтверждена с помощью метода меченых атомов в 1951 г. Кейнесом и др. Они показали, что во время возбуждения в нервное волокно действительно входит гораздо больше ионов Ма+, чем в покое. Оказалось, что и выход калия наружу тоже не перекращается, даже наоборот, становится сильнее, чем в покое.
Итак, основная идея, введенная Ходжкином и Хаксли в мембранную теорию, идея о том, что проницаемость мембраны для того или иного иона не является величиной постоянной, а может меняться в зависимости от условий, оказалась верной.
Но одной этой общей идеи для объяснения механизма возбуждения и всех явлений, связанных с ним, было явно недостаточно.
Пока не было установлено, как в действительности во время возбуждения меняется проницаемость мембраны для калия и для натрия и, главное, от каких условий зависят эти изменения.
Опыты с внутриклеточным раздражением аксона показывали, что для возникновения возбуждения необходимо изменить на определенную величину мембранный потенциал, довести его до порогового значения. Изменения порога в течение рефрактерного периода зависели от времени. Была известна зависимость «сила – длительность» и т.д. Поэтому Ходжкин и Хаксли предположили, что проницаемость мембраны определяется двумя факторами: мембранным потенциалом и временем, прошедшим с момента изменения потенциала. Надо было выяснить, верно ли это предположение.
А выяснить это было чрезвычайно трудно, и не только потому, что процесс возбуждения, как мы уже не раз отмечали, очень быстрый. Дело еще и в том, что всякое изменение потенциала на мембране – а в этом, по существу, и состоит явление возбуждения – вызывает, согласно гипотезе, изменение проницаемости и для натрия, и для калия, а изменение проницаемости сейчас же меняет потенциал. Существенно также, что всякое изменение потенциала сейчас же вызывает в мембране емкостный ток: ведь в электрическом отношении мембрана – это сопротивление и емкость, включенные параллельно.
Решающую роль в экспериментальных исследованиях проницаемости клеточной мембраны сыграла разработка Ходжкином и Хаксли так называемого метода фиксации потенциала. Вот где пригодился Ходжкину опыт, приобретенный во время войны в работе с радарными установками! Ходжкин сконструировал специальный прибор, который с помощью электронных схем улавливал малейшие изменения мембранного потенциала и очень быстро подавал на мембрану ток, компенсирующий эти изменения. С помощью такой управляющей схемы с обратной связью на мембране поддерживался постоянный потенциал., причем в течение любого нужного времени.
Теперь, можно сказать, руки были развязаны: во-первых, раз потенциал на мембране не меняется, ток через емкость не идет; во-вторых, зафиксировав потенциал, можно было проверить, действительно ли, как предполагалось, проницаемость меняется со временем уже сама по себе, независимо от потенциала.
Опыт ставился так. На мембране за очень короткое время устанавливали необходимую разность потенциалов и поддерживали ее на заданном уровне достаточно долго, регистрируя ток, пропускаемый через мембрану. При этом сила тока была пропорциональна проводимости *) мембраны в данный момент времени.
Чтобы разделить калиевую и натриевую проницаемости, использовали в качестве окружающей волокно среды раствор, содержащий только исследуемый ион, или специальные вещества, которые препятствовали прохождению другого иона через мембрану.
Проведя на своей установке обширную серию экспериментов, Ходжкин и Хаксли получили эмпирические
графики зависимостей изменения проницаемости от сдвига потенциала и времени, точнее, «срезы» этих функций для различных значений потенциала. Было показано, что каждый сдвиг МП от положения равновесия, т.е. от ПП, равного 80 мВ, действительно вызывает изменение проницаемости мембраны и для калия, и для натрия.
Результат одного из таких опытов по изучению калиевой проницаемости показан на рис. 19. Здесь по оси ординат отложен ток, проходящий через единицу поверхности мембраны, а по оси абсцисс – время, начиная с момента установления потенциала на заданном уровне.
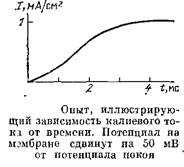
Видно, что ток постепенно нарастает, а это значит, что сопротивление мембраны снижается: при постоянном МП по закону Ома произведение тока на сопротивление постоянно. Но снижение сопротивления означает рост проводимости мембраны для ионов калия. Этот результат показывает, что калиевая проницаемость действительно зависит не только от МП, но и от времени.
Теперь мы попросим уважаемого читателя внимательно разобраться в полученных в результате таких экспериментов графиках, изображенных на рис. 20, а, б. На первом изображена зависимость изменения калиевой проводимости от сдвига потенциала и времени, на втором – натриевой. Если мембранный потенциал равен примерно –80 мВ, т.е. потенциалу покоя, то и калиевая, и натриевая проводимости не меняются, причем первая примерно в 25 раз больше, чем вторая, и равна примерно 1 мС/см2 *). Поэтому в нашем масштабе исходная проницаемость для калия почти не видна. При сдвиге мембранного потенциала, например, от –80 до –42 мВ, т.е. при деполяризации мембраны на 38 мВ, калиевая проводимость увеличивается, причем сначала очень медленно: первую миллисекунду почти не меняется, потом примерно за 5–8 мс постепенно увеличивается и через некоторое время устанавливается на новом постоянном уровне. На кривых рис. 20, а видно, что этот новый уровень тем выше, чем больше сдвиг потенциала на мембране, однако он не превосходит некоторого определенного значения.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики