Биохимия сахарного диабета
ППГ-белки могут гидролизоваться макрофагами (с участием ППГ-рецепторов) или межклеточными протеолитическими системами с образованием ППГ-пептидов, часто длиной около 30 аминокислотных остатков. ППГ-белки, и особенно образующиеся в результате их гидролиза ППГ-пептиды, попадают и в кровоток. Концентрация ППГ-пептидов в крови резко повышается при почечной недостаточности разного происхождения, в т
ом числе при диабетической нефропатии. Это связано с тем, что элиминация ППГ-пептидов поисходит с участием почек: ППГ-пептиды фильтруются в клубочках, реабсорбируются клетками проксимальных канальцев и катаболизируются в лизосомах этих клеток.
В экспериментах на крысах показано, что введение ППГ-белков в кровь приводит к ковалентному связыванию этих белков с белками межклеточного матрикса во многих тканях и к появлению структурных и функциональных нарушений, сходных с теми, которые бывают при сахарном диабете.
ППГ проявляют многообразную биологическую активность: повышают проницаемость эндотелиальных клеток, соединяются с рецепторами макрофагов, эндотелиальных и мезангиальных клеток, активируют макрофаги к секреции цитокинов (рецепторным путем), подавляют образование NO и соответственно ингибируют расширение сосудов, усиливают окисление ЛНП. В крови больных диабетом обнаруживаются антитела к ППГ-пептидам.
6. Диабетические ангиопатии
Первичные проявления ангиопатий связаны с повреждением базальных мембран сосудов. Базальные мембраны (БМ) представляют собой пленки, на которых “растут” все клетки организма, кроме клеток соединительной ткани и крови: по одну сторону располагается клетка или слой клеток, а другой стороной БМ контактирует с фиброретикулярным межклеточным матриксом:
|
Базальные мембраны разных органов (А) и капилляров почечного клубочка (Б): а – люмен капилляры; б – полость Боуменовой капсулы; 1 – эндотелий; 2,3,4 – БМ клубочка (2 – lamina rara interna, 3 – lamina densa, 4 – lamina rara externa); 5 – подоцит, отростками контактирующий с БМ. |
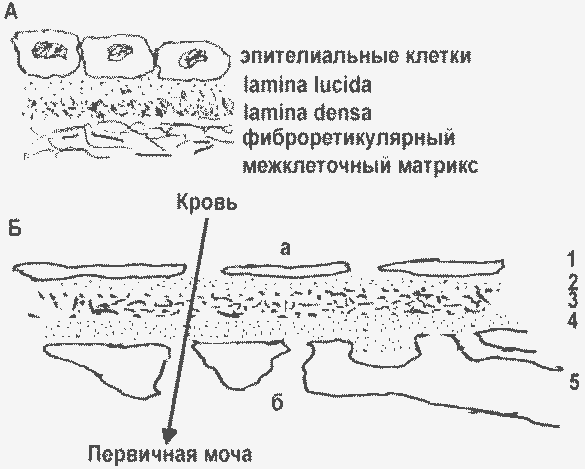
Эндотелий кровеносных сосудов, в том числе капилляров, тоже располагается на базальных мембранах. В отличие от всех прочих органов, в капиллярах почечного клубочка БМ трехслойна, а клетки располагаются по обе ее стороны.
В построении БМ участвуют коллагены, протеогликаны, неколлагеновые гликопротеины. Все компоненты БМ синтезируются прилегающими к ним клетками. Специальные функции выполняют интегрины - белки плазматической мембраны клеток, соединяющие клетку с БМ.
Коллаген IV типа - основной структурный белок базальных мембран. В геноме человека имеется шесть локусов, кодирующих шесть различающихся пептидных цепей, из которых строятся трехцепочечные молекулы коллагена IV. Чаще всего коллаген IV содержит цепи 1(IV) и 2(IV) в составе гетеротримеров [1(IV)]22(IV). Коллаген IV относится к сетеобразующим коллагенам. Взаимодействуя С-концевыми глобулярными доменами, молекулы образуют димеры, а при взаимодействии N-концевыми глобулярными доменами - тетрамеры.
Наряду с этими взаимодействиям конец в конец возможны и латеральные взаимодействия трехцепочечных спиральных доменов, в том числе с образованием суперспиралей. В конечном счете образуется сетевидная трехмерная структура с гексагональными ячейками размером 170 нм. Коллаген IV имеет также центры связывания с некоторыми белками клеточной мембраны, в том числе с интегринами.
Значительную часть массы БМ составляют протеогликаны. Эти молекулы содержат белковое ядро и ковалентно связанные с ним гликозамингликаны. В БМ в наибольших количествах содержатся гепарансульфат-протеогликаны (ГСПГ), и в значительно меньших - хондроитинсульфат-протеогликаны.
Гепарансульфат представляет собой неразветвленную цепь, построенную из глюкуроновой кислоты и глюкозамина, с последовательностью (ГлкА-ГлкN)n. Остаток глюкозамина может быть сульфирован по 2-й, 3-й и 6-й позициям. Молекулярная масса цепей обычно от 50 до 100 кДа. С одним белковым ядром ГСПГ обычно связано несколько цепей гепарансульфата. ГСПГ в БМ соединяется с коллагеном IV и ламинином определенными центрами белкового ядра и цепей гепарансульфата. Кроме того, ГСПГ разными способами может быть связан с поверхностью клеток, с участием как гликозамингликановой части, так и белкового ядра.
Ламинин - специфичный для БМ неколлагеновый гликопротеин. Молекула ламинина - тример , имеет крестообразную форму, с тремя короткими ветвями и одной длинной:
|
Молекулы базальных мембран. А. Многомолекулярные структуры, образуемые коллагеном IV: 1 - тетрамер; 2 - димер; 3 - мономер. Б. Фибронектин. В. Основные гепарансульфат-протеогликаны БМК: 1 - перлекан; 2 - ГСПГ 200 кДа; 3 - ГСПГ 350 кДа (горизонтальные линии - пептидные цепи, вертикальные линии - гепарансульфатные цепи). Г. Ламинин. |
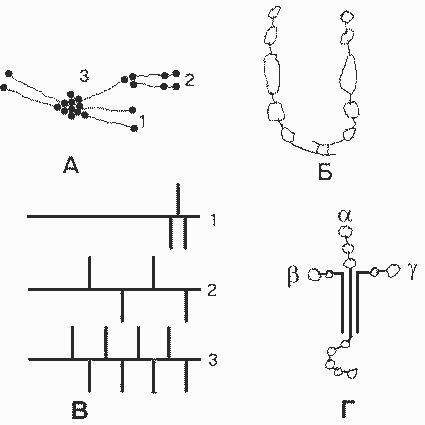
Известны три разные цепи , три цепи и две цепи . Варианты цепей могут комбинироваться по-разному при образовании тримерной молекулы. Пока обнаружено семь разных ламининов. Каждая из ветвей содержит глобулярные домены, которые имеют ряд центров свяэывания разных лигандов. Короткие ветви участвуют в образовании БМ путем взаимодейсв с другими молекулами ламинина, с коллагеном IV (при участии еще одного белка - нидогена), а также с интегрином 11 клеточной мембраны. Глобулярный домен длинной ветви участвует в межклеточной адгезии, взаимодействуя с разными интегринами и другими белками плазматической мембраны клеток. Длинная ветвь взаимодействует также с гепарансульфатными протеогликанами.
Интегрины представляют собой трансмембранные гликопротеины, -димеры. Каждая цепь пересекает мембрану один раз. Обе цепи имеют большие внеклеточные домены (аминоконцевые), образующие центр связывания, комплементарный соответствующему лиганду - компоненту матрикса. Внутриклеточный домен взаимодействуют с актиновым цитоскелетом при посредстве ряда промежуточных белков. С местом взаимодействия интегринов с цитоскелетом соседствуют сигнальные белки, которые активируются, когда к внеклеточному домену интегрина присоединяется лиганд. Таким лигандом могут быть ламинины, коллаген IV, протеогликаны.
Сила, с которой интегрин связывается с лигандом, может быстро и с высокой точностью регулироваться - свойство, которое называют “модуляция сродства”. В покоящемся (“неактивном”) состоянии интегрины имеют низкое сродство к своим лигандам, и это характерно для нормальных физиологических условий. Определенные стимулы превращают их в активные рецепторы с высоким сродством к лигандам. Это позволяет клеткам быстро приспосабливать их адгезивные свойства к изменившимся условиям без изменения количества молекул адгезии. А поскольку сигнал, полученный интегрином, передается в клетку, то могут изменяться не только адгезивные свойства, но и внутриклеточные процессы.
Другие рефераты на тему «Медицина»:
Поиск рефератов
Последние рефераты раздела
- Особенности лечения и тракционно-экстензионной терапии на аппарате Kinetrac KNX-7000
- Остеохондроз, методики лечения
- Тракционно-экстензионная терапия у больных остеохондрозом пояснично-крестцового отдела позвоночника
- Болезни, возникающие от курения. Профилактика курения
- Болезни органов дыхания
- Болезни желчевыводящих путей и печени
- Анатомия и физиология артерий нижних конечностей. Этиология и патогенез