Динамическое поведение мембран
6. Модели описывающие латеральную дифузию
Для описания поступательной диффузии мембранных белков часто применяется гидродинамическая модель Саффмана и Дельбрюка. В этой модели рассматривается диффузия белка в тонком вязком слое; при этом считается, что растворитель является сплошной средой, т. е. молекулы растворителя малы по сравнению с диффундирующими молекулами. В рамках этой моде
ли выполняется следующее соотношение:
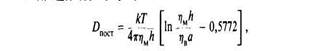
где и ηв — вязкости мембранной и водной фаз соответственно, а — радиус белковой молекулы, имеющей форму цилиндра, h — толщина мембраны. Из модели следует, что скорость поступательной диффузии слабо зависит от размера молекулы, что подтверждается полученными к настоящему времени экспериментальными данными (табл2.).
Таблица коэффициенты латеральной диффузии
A. В фосфодипидных везикулах
Липидные метки (ди1, NBD-PE) 1—5 100
в фосфолипидных везикулах
(см. табл. 5.1)
NBD-Грамицидин 2 100
Гликофорин 2 100
Родопсин 1—3 100
Ацетилхолиновый рецептор 1—2 100
Белок полосы 3 1—2 100
Б. В природных биомембранах
Липидные метки 0,2—2 100 Рецептор для фактора роста нервов
(нейриты) 0,06—0,08 22—64
Рецептор lgE (тучные клетки крысы) 0,03 50—80
Родопсин (диски палочек сетчатки) 0,4 100
Ацетилхолиновый рецептор
(миобласты) < 0,0003 — Белок полосы 3
(тени эритроцитов) 0,004 10
B. В мембранах при нарушении связи с элементами цитоскелета
Ацетилхолиновый рецептор (вздутия в
мембране моторной концевой
пластинки) 0,3 100
Белок полосы 3 (эритроцит,
дефицитный по спектрину) 0,2
Эта гидродинамическая модель не позволяет описывать диффузию липидных зондов, поскольку к ним неприменимо допущение о том, что белок диффундирует в сплошной среде. Для объяснения поведения липидов применялись различные модели, основанные на концепции свободного объема. В рамках этих моделей диффузия рассматривается как процесс движения молекул в полостях, образующихся вследствие спонтанных флуктуации, при этом определяющей характеристикой мембран является отношение площади, приходящейся на одну молекулу липида, к минимальной Удельной площади, соответствующей максимально плотной упаковке липидов, т. е. плотность упаковки липидов. Как было отмечено, скорость вращения флуоресцентных зондов зависит от плотности упаковки липидов в мембране. Применимость этой модели к описанию вращательной или поступательной диффузии липидов строго не доказана, поскольку изначально она была построена для неассоциированных жидкостей, к которым можно отнести далеко не все мембранные липидные структуры. Тем не менее обратная зависимость между скоростью поступательной диффузии липидов и плотностью упаковки липидов в мембранном бислое обычно существует.
Наконец, несколько подходов использовалось для теоретического объяснения влияния мембранных белков на латеральную диффузию белков и липидов в мембране.
Дело в том, что белки оказываются на пути диффундирующих молекул и вынуждают их перемещаться только в пространстве, не занятом белками (так называемый эффект архипелага. Обычно из-за этого коэффициенты поступательной диффузии в биомембранах оказываются примерно в 10 раз меньше (диффузия происходит медленнее), чем в чисто липидных бислоях.
7. Липидно-белковые взаимодействия
Большинство методов, применяемых для изучения упорядоченности и динамических свойств мембран, используется и для исследования липидно-белковых взаимодействий. Работы по изучению этих взаимодействий были в основном направлены на выяснение влияния мембранных белков на физическое состояние липидов. Рассмотрим типичную мембрану с весовым соотношением липид/белок = 1:1 (табл. 1.4). При средней мол. массе белка 50 кДа молярное соотношение липид/белок составляет ~ 60:1 при условии, что присутствуют только фосфолипиды. Для сравнения укажем, что соответствующие соотношения для мембраны наружного сегмента палочки сетчатки и саркоплазматического ретикулума составляют по оценкам 75:1 и 110:1. Если белок представляет собой цилиндр, выступающий за пределы бислоя с обеих сторон примерно на 10 А, то его радиус должен составлять около 18 А. Молекула фосфолипида в жидкой мембране занимает площадь ~60А2, что соответствует радиусу головки -4,4 А, и для того, чтобы полностью окружить белок, иа каждой стороне бислоя должно находиться около 16 таких молекул липидов. Следовательно, согласно этой модели, в любой момент времени около 50% липидов должно соседствовать с белковыми молекулами. Однако белком занято 35<Уо площади поверхности мембраны, и даже учитывая сугубо приближенный характер этой модели, можно понять, что физические методы регистрации состояния липидов должны учитывать влияние белков на свойства биомембран.
Чтобы выяснить структуру и функции мембран, необходимо прежде всего ответить на следующие вопросы: 1) насколько прочно связаны внутримембранные белки с липидами и какова природа ли-пидного слоя, прилегающего к белку? 2) как далеко простирается влияние мембранных белков на укладку и динамические свойства мембранных липидов? 3) как влияют липиды на структуру и функции внутримембранных белков? 4) как периферические мембранные белки, связанные с поверхностью бислоя, взаимодействуют с липидами и влияют на их поведение?
8. Скорость обмена пограничных липидов со свободными липидами
Термин «пограничный липидный слой» был введен в связи с появлением идеи о существовании стационарного (неподвижного) слоя липида, связанного с поверхностью белка, например цитохромоксидазы. Под «стационарностью» понимается отсутствие обмена липидов за время, сравнимое со временем оборота фермента, т. е. ~10~3с, поэтому такой фермент вместе с его непосредственным окружением может рассматриваться как долгоживущий комплекс. Во всех изученных к настоящему времени случаях это условие не выполняется, но термин «пограничный липидный слой» остается полезным описательным термином, если речь идет о ближайшем липидном окружении белка.
Данные 2Н-ЯМР четко показали, что пограничные липиды быстро обмениваются с основной массой липидов в бислое. Спектры ЭПР меченых фосфолипидов свидетельствуют о существовании свободных и связанных липидных компонентов, в то же время аналогичные эксперименты с дейтерированными липидными зондами гокомпонентности мембраны может быть оптимизация упаковки ли-пидов вокруг отдельных мембранных белков, уменьшающая возможные деформации иа границе белок—липид.
2. Другой тип деформаций, возникающих при встраивании белка в мембрану, — это латеральные искривления. Примером такого белка может служить «непрочно» связывающаяся форма цитохрома bs
3. Еще одной причиной деформаций бислоя может служить несоответствие между размером данного гидрофобного участка бислоя и толщиной мембраны. Чтобы избежать экспонирования гидрофобных областей в воду, белок или липиды могут частично изменить свою конформацию. Если белок не деформируется, то может произойти следующее: а) или ацильные цепи, или белковая молекула наклоняются относительно нормали к бислою на угол, зависящий от толщины мембраны. Это предположение было в одном случае подвергнуто проверке и не нашло подтверждения; б) ацильные цепи липидов деформируются; в) в гетерогенной смеси липидов последние реорганизуются таким образом, что молекулы «неправильной» длины оказываются сгруппированными вокруг белка.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики