Молекулярные механизмы многообразия антител
СОДЕРЖАНИЕ:
Введение
1. Строение антител
1.1 Функциональные свойства антител
1.2 Строение антител и их многообразие
1.3 Тонкая структура антител
2. Молекулярные механизмы многообразия антител
Список литературы
ВВЕДЕНИЕ
По имеющимся оценкам, у мыши может вырабатываться от 106 до 1012 различных молекул антител, совокупность которых называ
ют репертуаром. Этот репертуар, видимо, достаточно велик для того, чтобы почти для каждой антигенной детерминанты нашелся подходящий антиген-связывающий участок. Поскольку антитела представляют собой белки, а белки копируются генами, способность животного производить миллионы разных антител представляет собой чрезвычайно сложную генетическую проблему: как синтезировать миллиарды разных белков, не привлекая к этому чрезмерно большого числа генов. В решении этой проблемы участвует ряд уникальных вне генетических механизмов.
1. СТРОЕНИЕ АНТИТЕЛ
Иммунная система выработалась в процессе эволюции позвоночных для защиты от инфекций. Она состоит из миллиардов лимфоцитов и включает миллионы различных клонов. Лимфоциты каждого клона несут на своей поверхности рецептор, который позволяет им связывать ту или иную «антигенную детерминанту» - определенную группировку в молекуле антигена. Существуют два класса лимфоцитов: В-клетки, вырабатывающие антитела, и Т-клетки, которые осуществляют иммунные реакции клеточного типа.
Уже на ранних стадиях своего развития В- и Т-клетки с рецепторами для антигенных детерминант молекул собственного организма элиминируются или супрессируются; в результате иммунная система в норме способна отвечать только на чужеродные антигены. Связывание чужеродного антигена с лимфоцитом вызывает иммунный ответ, направленный против этого антигена. При этом некоторые из лимфоцитов пролиферируют и дифференцируются в клетки памяти, так что при вторичном воздействии того же антигена иммунный ответ развивается быстрее и оказывается гораздо более сильным.
1.1 Функциональные свойства антител
Единственная известная функция В-лимфоцитов – выработка антител. Уникальная особенность антител, отличающая их от всех других известных белков, состоит в том, что они могут существовать в миллионах разновидностей – каждая со своим уникальным участком для связывания антигена. В совокупности называемые иммуноглобулинами(сокращенно Ig) антитела образуют один из основных классов белков крови, составляя по весу примерно 20% суммарного белка плазмы.
Как и предсказывала гипотеза клональной селекции, все молекулы антител, производимые какой-то одной В-клеткой, имеют одинаковый антиген-связывающий участок. Первые антитела, синтезированные вновь образовавшейся В-клеткой, не секретируются; вместо этого они встраиваются в плазматическую мембрану, где служат рецепторами для антигена. Каждая В-клетка имеет на своей плазматической мембране приблизительно 105 таких молекул. Когда антиген присоединяется к молекулам антител на поверхности покоящейся В-клетки, это обычно инициирует сложную и малоизученную цепь событий, приводящую к клеточной пролиферации и дифференцировке с образованием клеток, секретирующих антитела. Такие клетки, вырабатывают большие количества растворимых (не связанных с мембраной) антител с таким же антиген-связывающим участком, что и у антител на поверхности клеток, и выделяют эти антитела в кровь. Активированные В-клетки могут начать секретировать антитела, будучи еще малыми лимфоцитами; конечная стадия этого пути дифференцировки - большая плазматическая клетка, которая выделяет антитела со скоростью около 2000 молекул в секунду. По-видимому, плазматические клетки используют для производства антител столь значительную часть мощности своего белоксинтезирующего аппарата, что не способны к дальнейшему росту и делению и погибают после нескольких дней секреции антител.
Простейшие молекулы антител имеют форму буквы Y с двумя идентичными антиген-связывающими участками – по одному на конце каждой из двух «ветвей» (рис. 1). Поскольку таких участков два, эти антитела называют валентными. Такие антитела могут сшивать молекулы антигена в обширную сеть, если каждая молекула антигена имеет три или большее число антигенных детерминант. Достигнув определенных размеров, такая сеть выпадает из раствора. Как мы увидим позже, тенденция больших иммунных комплексов к осаждению (преципитации) удобна для выявления антител и антигенов. Эффективность реакций связывания и сшивания антигена антителами значительно возрастает благодаря гибкому шарнирному участку в месте соединения обеих «ветвей» с «хвостом»: этот участок позволяет изменять расстояние между двумя антиген-связывающими участками (рис. 2). Защитное действие антител объясняется не просто их способностью связывать антиген. Они выполняют и целый ряд других функций, в которых участвует «хвост». Эта область молекулы определяет, что произойдет с антигеном, если он оказался связанным. Антитела с одинаковыми антиген-связывающими участками могут иметь весьма разные «хвостовые» области, а потому и разные функциональные свойства.
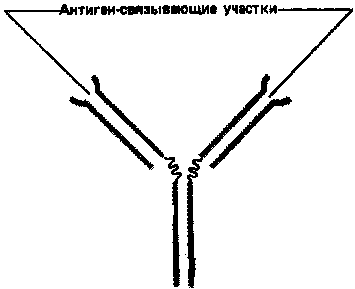
Рисунок 1. Сильно упрощенная схема молекулы антитела с двумя идентичными антиген-связывающими участками.
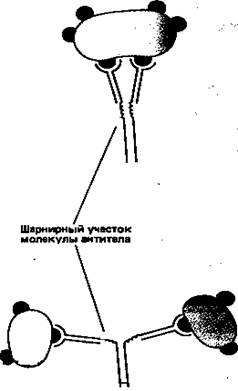
Рисунок 2. Шарнирный участок молекулы антитела повышает эффективность связывания молекул антигена и сшивания их друг с другом.
1.2 Строение антител и их многообразие
Основную структурную единицу молекулы антитела образуют четыре полипептидные цепи – две идентичные легкие (L-цепи, каждая примерно из 220 аминокислот) и две идентичные тяжелые (Н-цепи, каждая примерно из 440 аминокислот). Все четыре цепи соединены между собой с помощью нековалентных взаимодействий и ковалентных связей (дисульфидных мостиков). Молекула состоит из двух одинаковых половинок, в которых L- и Н-цепи вносят почти равный вклад в построение двух идентичных антиген-связывающих участков (рис. 3).
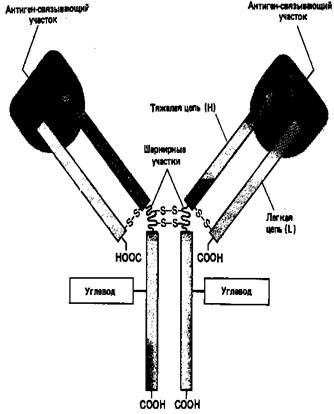
Рисунок 3. Схематическое изображение типичной молекулы антителу состоящей из двух идентичных тяжелых (Н) и двух идентичных лёгких (L) цепей. Антиген-связывающие участки формируются за счет комплекса N-концевых областей L- и Н-цепей, а область «хвоста» образуют только Н-цепи. Каждая цепь содержит одну или несколько олигосахаридных цепочек, функция которых не известна.
Протеолитические ферменты папаин и пепсин расщепляют молекулы антител на различные характерные фрагменты: папаин дает два отдельных идентичных Fab-фрагмента, каждый из которых обладает одним антиген- связывающим участком, и один Fc-фрагмент (Fab - сокращение слов fragment antigen binding; Fc означает «кристаллизующийся фрагмент» (от crystallizable).; пепсин дает один Р(аb')2-фрагмент, состоящий из двух ковалентно связанных Р(аb')-фрагмент (каждый из которых немного больше, чем Fab-фрагмент), и много более лёгких фрагментов (рис. 4). Поскольку Р(аb')2-фрагменты бивалентны в отличие от моновалентных Fab-фрагментов сохраняют способность связывать антигены и образовывать преципитаты. Ни один из этих фрагмент обладает другими биологическими свойствами нативных молекул антигенов поскольку они не содержат «хвостовой» (Fc) области, определяющей их свойства.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики