Компьютерное моделирование в экологии
Кроме минимального содержания элемента qmin в клетке, важной характеристикой вида является величина qmax – максимальное значение элемента питания, которое может накопиться в клетке. Можно привести несколько примеров использования величины qmax в формулах для скорости роста клеток в модели весеннего цветения диатомовых на Балтике применял зависимость

А.Дота в модели поликультуры пресноводных водорослей использовал выражение
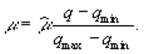
На непостоянство стехиометрических соотношений углерода, азота и фосфора в составе фитопланктона указывал С.Йоргенсен (1985). Он отмечал, что, как показали лабораторные опыты, развитие фитопланктона происходит в два этапа: на первом идет поглощение биогенных веществ, а на втором – клеточное деление, регулируемое внутриклеточной концентрацией ресурсов. С.Йоргенсен (Jørgensen, 1976) использует следующую систему уравнений
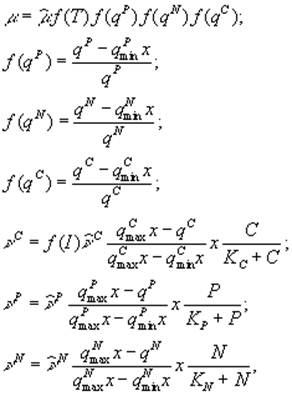
где ![]() – внутриклеточные концентрации биогенных элементов (соответственно, фосфора, азота и углерода), выраженные в мг на 1 л воды;
– внутриклеточные концентрации биогенных элементов (соответственно, фосфора, азота и углерода), выраженные в мг на 1 л воды; ![]() – минимальные и максимальные значения содержания трех биогенных веществ в составе фитопланктона; x – концентрация фитопланктона;
– минимальные и максимальные значения содержания трех биогенных веществ в составе фитопланктона; x – концентрация фитопланктона; ![]() – скорости поглощения углерода, фосфора, азота;
– скорости поглощения углерода, фосфора, азота; ![]() – соответствующие максимальные скорости поглощения; C, P, N – концентрации биогенных элементов в окружающей среде;
– соответствующие максимальные скорости поглощения; C, P, N – концентрации биогенных элементов в окружающей среде; ![]() – функции, характеризующие зависимости интенсивности фотосинтеза от освещенности и температуры.
– функции, характеризующие зависимости интенсивности фотосинтеза от освещенности и температуры.
Существуют некоторые данные, которые позволяют сравнить результаты моделирования с использованием концепции клеточной квоты и без ее использования. Л.А.Кучай (Кучай, 1985) специально сравнивала модели, описывающие кругооборот биогенных ресурсов (азота или азота и фосфора) в замкнутой системе, включающей одну группу фитопланктона, питательные элементы и детрит. При использовании клеточной квоты удельная скорость роста задавалась формулой Друпа, при этом в случае двух ресурсов использовался принцип минимума Либиха. Оказалось, что модели с использованием клеточной квоты дают большее число возможных сценариев развития фитопланктона и позволяют описывать ситуации, реально встречающиеся в природе, например, ситуацию нарастания биомассы фитопланктона в условиях почти нулевой концентрации биогенного элемента в среде или ситуацию резкого скачка отношения N:P в среде в момент кульминации цветения.
В настоящее время среди исследователей установилось мнение, что модель Моно адекватна при описании устойчивого роста водорослей в стационарных условиях, подобных хемостату, а для описания процесса роста при изменяющихся внешних условиях, например, при моделировании внутригодовой сукцессии фитопланктона, необходима более сложная модель, выбранная в соответствии с предположением зависимости скорости роста от клеточных концентраций ресурсов. Одной из ситуаций, где при моделировании необходимо привлечение концепции клеточной квоты, является накопительное культивирование.[1]
Дифференциальные уравнения в микробиологии. Дифференциальные уравнения, описывающие и концентрации микробной биомассы (x) и концентрации лимитирующего субстрата (s) в условиях хемостатного культивирования, которые были выведены Ж.Моно исходя из условий материального баланса, составили первую модель роста микробных популяций.
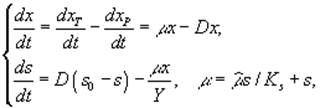
где xP – концентрация микроорганизмов в приемнике, куда сливается нарастающая бактериальная суспензия; xT – суммарная концентрация микроорганизмов (xT = x + xP); μ – удельная скорость роста, ![]() – максимальная скорость роста; s – концентрация лимитирующего ресурса в среде; Ks – константа полунасыщения при лимитировании данным субстратом; s0 – величина концентрации лимитирующего субстрата на входе в культиватор; D – скорость разбавления, равная отношению скорости поступления питательной среды к объему культуры; Y – экономический коэффициент (выход биомассы на единицу потребленного субстрата).
– максимальная скорость роста; s – концентрация лимитирующего ресурса в среде; Ks – константа полунасыщения при лимитировании данным субстратом; s0 – величина концентрации лимитирующего субстрата на входе в культиватор; D – скорость разбавления, равная отношению скорости поступления питательной среды к объему культуры; Y – экономический коэффициент (выход биомассы на единицу потребленного субстрата).
Принципиальной особенностью данной открытой системы является возможность установления динамического равновесия. В установившемся состоянии μ = D, а ![]() (
(![]() – постоянные значения стационарного состояния).[2]
– постоянные значения стационарного состояния).[2]
Необходимо также отметить, что при превышении скорости разбавления D критического значения 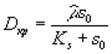 наступает режим вымывания, т.е. скорость потока настолько велика, что прирост биомассы не может компенсировать ее отток. Нулевое значение концентрации биомассы и равенство концентрации лимитирующего ресурса в среде значению последней на входе в культиватор является неустойчивым положением равновесия при скорости разбавления ниже критической и устойчивым при больших скоростях
наступает режим вымывания, т.е. скорость потока настолько велика, что прирост биомассы не может компенсировать ее отток. Нулевое значение концентрации биомассы и равенство концентрации лимитирующего ресурса в среде значению последней на входе в культиватор является неустойчивым положением равновесия при скорости разбавления ниже критической и устойчивым при больших скоростях ![]() .
.
Для различных методов проточного культивирования динамика концентрации биомассы микроорганизмов описывается соответствующими дифференциальными уравнениями (Паников, 1991).
1) Турбидостат 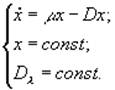
![]() – оптическая плотность.
– оптическая плотность.
2) pH-стат 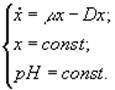
3) Хемостат с возвратом 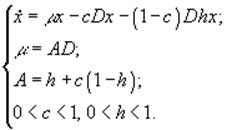
4) Батарея хемостатов 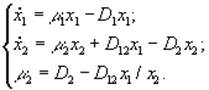
Наряду с различными способами гомогенного непрерывного культивирования для лабораторных исследований применяют и другие методы:
1) Непрерывное культивирование без выноса микробных клеток:
а) периодический диализ 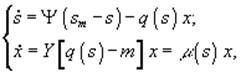
б) непрерывный диализ 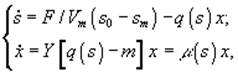
в) периодическая культура с подпиткой (ПКП)
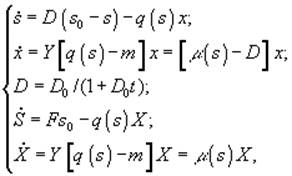
г) периодическая культура с газовым питанием
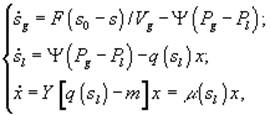
![]() – величины концентрации лимитирующего субстрата в диализном сосуде, в газовой и жидкой фазах; Pg и Pl – парциальные давления; Ψ – коэффициент массопереноса;
– величины концентрации лимитирующего субстрата в диализном сосуде, в газовой и жидкой фазах; Pg и Pl – парциальные давления; Ψ – коэффициент массопереноса; ![]() – объемы диализного сосуда, воздушной фазы ферментера,; S, X – количество субстрата и биомассы в расчете на всю культуру (s, x – в расчете на единицу объема); F – поток лимитирующего субстрата; s0 – исходная концентрация лимитирующего субстрата; q(s) - скорость потребления лимитирующего субстрата.[3]
– объемы диализного сосуда, воздушной фазы ферментера,; S, X – количество субстрата и биомассы в расчете на всю культуру (s, x – в расчете на единицу объема); F – поток лимитирующего субстрата; s0 – исходная концентрация лимитирующего субстрата; q(s) - скорость потребления лимитирующего субстрата.[3]
Другие рефераты на тему «Экология и охрана природы»:
Поиск рефератов
Последние рефераты раздела
- Влияние Чекмагушевского молочного завода на загрязнение вод реки Чебекей
- Влияние антропогенного фактора на загрязнение реки Ляля
- Киотский протокол - как механизм регулирования глобальных экологических проблем на международном уровне
- Лицензирование природопользования, деятельности в области охраны окружающей среды и обеспечения экологической безопасности
- Мировые тенденции развития ядерной технологии
- Негативные изменения состояния водного бассейна крупного города под влиянием деятельности человека
- Общественная экологическая экспертиза и экологический контроль