Компьютерное моделирование в экологии
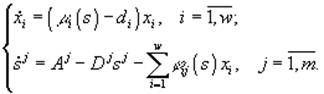 (1)
(1)
Модель, описывающая культивируемое в хемостате сообщество, является частным случаем модели (3.1) при условиях di = Dj = D, ![]() = Aj/D, где D – скорость разведения культуры,
= Aj/D, где D – скорость разведения культуры, ![]() – концентрация j-го субс
– концентрация j-го субс
трата в поступающей в реактор питательной среде, и задается системой уравнений
![]()
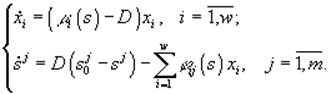
(Хемостат – непрерывный культиватор, в котором за счет саморегулирующх механизмов популяции поддерживается постоянная плотность популяции.)
В случае взаимозаменяемых ресурсов зависимость i(s) описывается выражением
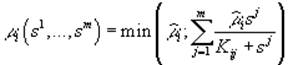
или его линейной аппроксимацией
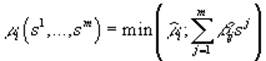 ,
,
где ij – элементы матрицы приспособленности: 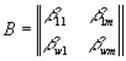 , здесь ij = cijYij, где cij – удельная скорость потребления j-го субстрата i-м видом организма, а Yij – экономический коэффициент использования j-го вещества i-м видом.
, здесь ij = cijYij, где cij – удельная скорость потребления j-го субстрата i-м видом организма, а Yij – экономический коэффициент использования j-го вещества i-м видом.
Если ресурсы взаимонезаменяемы, то
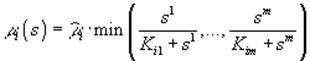
или
![]() .
.
В приведенных выше формулах символ ![]() обозначает максимально возможную удельную скорость роста организмов i-го вида; Kij – константа полунасыщения роста i-го вида на j-м субстрате.
обозначает максимально возможную удельную скорость роста организмов i-го вида; Kij – константа полунасыщения роста i-го вида на j-м субстрате.
В моделях, использующих дифференциальные уравнения, возможен учет лимитирования роста световой энергией. В этом случае предлагается использовать следующую зависимость:
![]()
здесь xi – концентрация клеток i-го вида; μi – удельная скорость роста i-го вида; ![]() – максимально возможная удельная скорость роста организмов i-го вида;
– максимально возможная удельная скорость роста организмов i-го вида; ![]() – процентное содержание хлорофилла в биомассе; li – коэффициент поглощения света i-м видом;
– процентное содержание хлорофилла в биомассе; li – коэффициент поглощения света i-м видом; ![]() – КПД фотосинтеза i-го вида клеток; Yi – величина, обратная калорийности биомассы i-го вида клеток; E – мощность ФАР (фотосинтетически активной радиации), падающей на поверхность культуры; d – толщина слоя культуры в кювете культиватора (приведено выражение для удельной скорости μi в случае лимитирования совместного роста двух видов только энергией света) (Абросов, Ковров, 1977).
– КПД фотосинтеза i-го вида клеток; Yi – величина, обратная калорийности биомассы i-го вида клеток; E – мощность ФАР (фотосинтетически активной радиации), падающей на поверхность культуры; d – толщина слоя культуры в кювете культиватора (приведено выражение для удельной скорости μi в случае лимитирования совместного роста двух видов только энергией света) (Абросов, Ковров, 1977).
А.Я.Болсуновским (1999) была предложена следующая математическая модель динамики компонентов сообщества из двух видов (спирулина и хлорелла), рост которых лимитируется интенсивностью света всей области ФАР
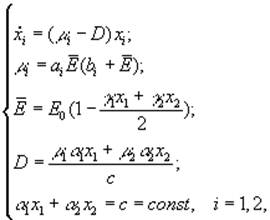
где xi – концентрация клеточного хлорофилла; μi – удельные скорости роста; ai, bi – константы уравнения Михаэлиса-Ментен; ![]() – средняя освещенность для всего сообщества; E0 – поверхностная освещенность; D – скорость протока.
– средняя освещенность для всего сообщества; E0 – поверхностная освещенность; D – скорость протока.
Кроме того, в моделях возможно отразить эффекты метаболического воздействия, как, например, это было сделано Ю.А.Домбровским с соавторами (1990). Скорость роста фитопланктона i -го вида описывалась выражением
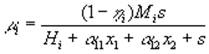 ,
,
где x1, x2, s – концентрации двух видов фитопланктона и минерального вещества, выраженные в единицах лимитирующего биогенного вещества; ![]() – коэффициенты метаболизма; Mi – максимальная скорость фотосинтеза; Hi – параметр насыщения;
– коэффициенты метаболизма; Mi – максимальная скорость фотосинтеза; Hi – параметр насыщения; ![]() – эмпирический коэффициент ингибирования i-го вида j-м.
– эмпирический коэффициент ингибирования i-го вида j-м.
Как уже указывалось выше, при моделировании динамики биомассы фитопланктона, чаще всего используют уравнение Моно, описывающее зависимость удельной скорости роста популяции μ от концентрации s лимитирующего рост субстрата
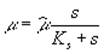 , (2)
, (2)
где ![]() – максимальная скорость роста; Ks – константа полунасыщения при лимитировании данным субстратом. (В общем случае
– максимальная скорость роста; Ks – константа полунасыщения при лимитировании данным субстратом. (В общем случае ![]() и Ks зависят от вида фитопланктона.) Теоретическое объяснение указанной зависимости обычно основывается на представлениях биохимической кинетики, связанных с концепцией “узкого места” метаболизма (Алексеев и др., 1992). Живая клетка представляет собой строго сбалансированную систему ферментативных реакций, при этом общая скорость прироста биомассы
и Ks зависят от вида фитопланктона.) Теоретическое объяснение указанной зависимости обычно основывается на представлениях биохимической кинетики, связанных с концепцией “узкого места” метаболизма (Алексеев и др., 1992). Живая клетка представляет собой строго сбалансированную систему ферментативных реакций, при этом общая скорость прироста биомассы ![]() предполагается пропорциональной скорости реакции, которая ограничена недостатком субстрата в среде. В простейшем случае зависимость скорости ферментативной реакции от концентрации субстрата описывается формулой Михаэлиса-Ментен. Поскольку в формуле Михаэлиса-Ментен константы
предполагается пропорциональной скорости реакции, которая ограничена недостатком субстрата в среде. В простейшем случае зависимость скорости ферментативной реакции от концентрации субстрата описывается формулой Михаэлиса-Ментен. Поскольку в формуле Михаэлиса-Ментен константы ![]() и Ks есть функции от скоростей прямой и обратной ферментативной реакции, то постоянство
и Ks есть функции от скоростей прямой и обратной ферментативной реакции, то постоянство ![]() и Ks предполагает постоянство этих скоростей. Таким образом, в основе использования зависимости Моно лежит неявное предположение об инвариантности во времени соотношения элементов в клетке (так как только при постоянстве состава клетки могут оставаться постоянными скорости ферментативных реакций). Кроме того, при подходе, основанном на зависимости Моно, процессы поглощения питательных веществ и роста фитопланктона не различаются.
и Ks предполагает постоянство этих скоростей. Таким образом, в основе использования зависимости Моно лежит неявное предположение об инвариантности во времени соотношения элементов в клетке (так как только при постоянстве состава клетки могут оставаться постоянными скорости ферментативных реакций). Кроме того, при подходе, основанном на зависимости Моно, процессы поглощения питательных веществ и роста фитопланктона не различаются.
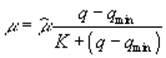 . (3)
. (3)
Формула (3.3) упрощается в связи с предположением, подтвержденным экспериментально, что константа полунасыщения и минимальное клеточное содержание соответствующего биогена равны, и принимает вид
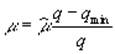 .
.
Дж.Фухсом в эксперименте на двух видах диатомовых продемонстрирована связь скорости роста водорослей с клеточной концентрацией фосфора в виде ![]() (Обозначения те же, что и в формулах (2), (3).)
(Обозначения те же, что и в формулах (2), (3).)
В.Бьерман (Bierman, 1976) в модели внутригодовой сукцессии фитопланктона (четыре группы водорослей, три биогенных элемента – азот, фосфор, кремний) выражал зависимость роста от клеточного фосфора формулой 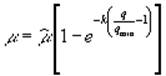 , от клеточного азота – формулой Друпа (3), а от клеточного кремния – формулой Моно (2).
, от клеточного азота – формулой Друпа (3), а от клеточного кремния – формулой Моно (2).
Другие рефераты на тему «Экология и охрана природы»:
Поиск рефератов
Последние рефераты раздела
- Влияние Чекмагушевского молочного завода на загрязнение вод реки Чебекей
- Влияние антропогенного фактора на загрязнение реки Ляля
- Киотский протокол - как механизм регулирования глобальных экологических проблем на международном уровне
- Лицензирование природопользования, деятельности в области охраны окружающей среды и обеспечения экологической безопасности
- Мировые тенденции развития ядерной технологии
- Негативные изменения состояния водного бассейна крупного города под влиянием деятельности человека
- Общественная экологическая экспертиза и экологический контроль