Особенности пропионовокислого брожения - применение пропионовокислых бактерий
Введение: общая информация о пропионовокислых бактериях
Пропионовокислые бактерии - неспороносные грамположительные неподвижные палочки размером 0,5—0,8 или 1,0—1,5 мкм (в молодых культурах — искривленные, слегка ветвящиеся палочки, в более старых — кокковидной формы). Образуют колонии жёлтого, оранжевого или красного цвета, растут как в аэробных, так и в анаэробных условиях.
П
ропионовокислые бактерии родственны по ряду свойств гетероферментативным молочнокислым бактериям Они, как и молочнокислые бактерии, не встречаются в почве или водоемах. Обитают в основном в рубце и кишечнике жвачных животных, в молочных продуктах (не в молоке). Пропионовокислые бактерии — возбудители пропионовокислого брожения, сбраживают глюкозу, лактозу и др. углеводы, а также некоторые спирты с образованием пропионовой и уксусной кислот и CO2. После молочнокислого брожения, когда лактоза превращена в молочную кислоту, начинают размножаться.
Пропионовокислые бактерии применяют для микробиологического синтеза витамина B12. Пропионовые бактерии могут синтезировать гемсодержащие белки. В их клетках обнаружены цитохромы.
Конструктивный метаболизм пропионовых бактерий претерпел дальнейшую эволюцию в сторону большей независимости от органических соединений внешней среды. Пропионовые бактерии характеризуются хорошо развитыми биосинтетическими способностями и могут расти на простой синтетической среде с аммонийным азотом в качестве единственного источника азота при добавлении к среде пантотеновой кислоты и биотина, а для некоторых видов и тиамина. У ряда пропионовых бактерий обнаружена способность к азотфиксации.
Местообитание пропионовых бактерий — кишечный тракт жвачных животных, молоко, твердые сыры, в приготовлении которых они принимают участие.
1. Пропионовокислое брожение: химизм и особенности
Основные продукты пропионовокислого брожения, вызываемого несколькими видами бактерий из рода Propionibacterium, — пропионовая (CH3CH2OH) и уксусная кислоты и CO2. Химизм пропионовокислого брожения сильно изменяется в зависимости от условий. Это, по-видимому, объясняется способностью пропионовых бактерий перестраивать обмен веществ, например, в зависимости от аэрации. При доступе кислорода они ведут окислительный процесс, а в его отсутствии расщепляют гексозы путём брожения. Пропионовые бактерии способны фиксировать CO2, при этом из пировиноградной кислоты и CO2 образуется щавелевоуксусная кислота, превращающаяся в янтарную кислоту, из которой декарбоксилированием образуется пропионовая кислота.
Основное энергетическое значение для пропионовокислых бактерий имеют так называемые ключевые реакции пропионовокислого брожения.
Под пропионовокислым брожением подразумевают биохимический процесс превращения бактериями сахара, молочную кислоту и ее солей в пропионовую кислоту. В этом брожении, кроме пропионовой кислоты, образуются и такие продукты, как уксусная кислота, углекислый газ, янтарная кислота, ацетоин, диацетил, другие летучие ароматические соединения - диметилсульфид, ацетальдегид, пропионовый альдегид, этанол и пропанол. Химизм данного брожения подобен типичному молочнокислому брожению с той разницей, что образовавшаяся молочная кислота в этом брожении не конечный продукт, а промежуточный. От других типов брожения пропионовокислое отличается высоким выходом АТФ, участием некоторых уникальных ферментов и реакций.
Пропионовокислым бактериям свойственен бродильный тип метаболизма: они расщепляют сахара по пути Эмбдена–Мейергоффа до пропионата, ацетата, СО2 и сукцината. Химизм пропионовокислого брожения хорошо изучен и описан.
В пропионовокислом брожении мы имеем дело с карбоксилированием пирувата, приводящим к возникновению нового акцептора водорода — ЩУК. Восстановление пировиноградной кислоты в пропионовую у пропионовокислых бактерий протекает следующим образом. Пировиноградная кислота карбоксилируется в реакции, катализируемой биотинзависимым ферментом, у которого биотин выполняет функцию переносчика CO2. Донором CO2-группы служит метилмалонил-КоА. В результате реакции транскарбоксилирования образуются ЩУК и пропионил-КоА:
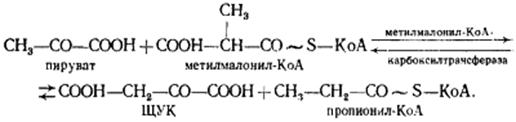
Ключевую реакцию брожения - превращение а-метилмалонил-КоА в сукцинил-КоА катализирует кофермент В12 (Ado Cbl).
Когда сбраживаемым субстратом является лактат, он сначала окисляется в пируват. Часть пирувата далее окисляется до ацетил-КоА и СО2, причем превращение ацетил-КоА в ацетат сопровождается образованием АТФ. Получение в процессе брожения окисленных продуктов, ацетата и СО2 уравновешивается сопутствующим восстановлением пирувата до пропионата.
Пировиноградная кислота - обязательное промежуточное соединение в брожении.
Пируват может быть превращен в пропионат несколькими путями:
1). пируват →акрилат→пропионат;
2). пируват→лактат→пропионат;
3). пируват + С 1 →сукцинат →метилмалонат →пропионат.
Первые две возможности у пропионовых бактерий не реализуются, и образование пропионата происходит из дикарбоновой кислоты по третьему пути.
Сначала лактат окисляется до пирувата при участии флавопротеида в качестве акцептора водорода, затем в реакции транскарбоксилирования образуется оксалоацетат. Донором СО2 служит (S)-метилмалонил-КоА, а переносчиком СО2 - биотин. Под действием малатдегидрогеназы и фумаразы образуется фумарат, который восстанавливается до сукцината в реакции, катализирующей фумаратредуктазой. Эта реакция сопряжена с синтезом АТР путем фосфорилирования, сопряженного с переносом электронов. Далее в КоА - трансферазной реакции образуется сукцинал-КоА. Затем под действием метилмалонил-КоА-мутазы, содержащей кофермент В12, осуществляется перегруппировка, ведущая к образованию (R)-метилмалонил-КоА, который, однако, не является субстратом для транскарбоксилазы. Скорее всего, (S)-стереоизомер образуется при действии специфической рацемазы. В зтом случае в реакции транскарбоксилирования синтезируется пропионил-КоА, и в результате последующего переноса КоА на сукцинат образуется пропионат.
В процессе сбраживания лактата в пропионат потребляется одна молекула NADH2. Она образуется при окислении лактата до ацетата в соответствии с суммарным уравнением реакций брожения:
Лактат + NADH2 + ADP + Pi → Пропионат + NAD + ATP.
Реакции, ведущие к образованию пропионовой кислоты у пропионовокислых бактерий, могут быть представлены следующей последовательностью:
1) Пируват + метилмалонил-КоА (а) ↔оксалоацетат+пропионил-КоА
2) Оксалоацетат+пропионил-КоА
3) Сукцинат + пропионил-КоА (+ КоА-трансфераза)↔сукцинил-КоА+пропионат
4) Сукцинил-КоА ↔(+метилмалонилизомеразая)↔метилмалонил-КоА
5) Метилмалонил-КоА (в) ↔(+метилмалонилрацемаза)↔метилмалонил-КоА(а)
Суммарно: пируват + 4H+→ пропионат.
Превращение оксалоацетата в сукцинат происходит в результате работы ферментов ЦТК: малатдегидрогеназы, фумаразы и сукцинатдегидрогеназы. Уксусная кислота образуется в результате окислительного декарбоксилирования пирувата:
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики