Особенности пропионовокислого брожения - применение пропионовокислых бактерий
6) Пируват + HAD+ + КоА(+пируватдегидрогеназа)↔ацетил-КоА+Н+ + НADH + CO2,
7) Ацетил-КоА + Фн (+фосфотрансацетилаза)↔ ацетил-Ф + КоА, ацетил-Ф + АDФ (+ацетилкиназа)↔ацетат + АТФ.
Итак, разобранный выше поток реакций приводит к синтезу пропионовой кислоты. Однако пропионовокислое брожение — более сложный процесс, поскольку наряду с пропионовой кислотой в качестве
продуктов брожения образуются уксусная, янтарная кислоты и CO2 (см. рис. 1):
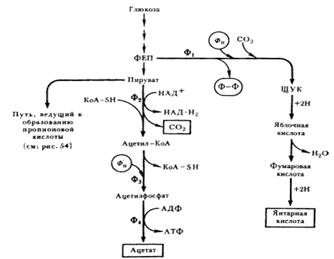
Рис. 1. Пути образования янтарной, уксусной кислот и CO2 пропионовыми бактериями: Ф1 — ФЕП-карбокситрансфосфорилаза; Ф2 — пируватдегидрогеназа; Ф3 — фосфотрансацетилаза; Ф4 — ацетаткиназа
Кроме основных продуктов в разных количествах в культуральной жидкости пропионовых бактерий обнаружены молочная, муравьиная, изовалериановая кислоты, этиловый и пропиловый спирты, уксусный и пропионовый альдегиды, ацетоин, диацетил. Состав конечных продуктов брожения зависит от культуры бактерий, состава среды и условий культивирования. Это касается как видов накапливаемых продуктов, так и количественных соотношений между ними.
Соотношение продуктов брожения может быть разное и в значительной степени зависит от степени окисленности источника углерода. При росте на среде с глицерином, например, отношение пропионовая: уксусная 2:1, с лактатом 1:1,5 и пируватом 1:2. С другой стороны, при культивировании пропионовокислых бактерий в строго анаэробных условиях соотношение между количествами кислот отклоняется в обратную сторону; именно в этом случае на одну молекулу уксусной всегда образуется 3 молекулы пропионовой. Также большое влияние имеет значение концентрации ионов водорода. При увеличении концентрации ионов водорода в среде изменяется соотношение основных продуктов брожения: образование уксусной кислоты увеличивается, а пропионовой заметно уменьшается. Соотношение пропионовой и уксусной кислот зависит от состава и свойств среды и внешних условий существования микроорганизмов. В сырах в период максимального развития культуры пропионовых бактерий (первая фаза) в основном образуются относительно окисленные соединения (уксусная кислота), в период спада развития – преимущественно более восстановленные (вторая фаза). Но при замедлении развития культуры P. shermanii (замена пептона аммонийными солями) уксусная кислота превалирует перед пропионовой, однако и в этом случае отношение пропионовой кислоты к уксусной возрастает. Этот пример служит доказательством связи продуцирования кислот с составом среды. Отношение пропионовой кислоты к уксусной зависит также от вида бактерий. В среде с глюкозой и дрожжевым автолизатом пропионовокислые бактерии рода P. thonii продуцируют указанные кислоты в отношении 5:1, для P. rubrum это отношение равно 3:1 и для P. shermanii – 2:1. Для культур P. shermanii (9 штаммов) отношения пропионовой кислоты к уксусной колеблется от 1,4 до 2,8.
Между количеством организмов и количеством образующихся кислот нет прямой связи. Изменение состава карбоновых кислот в питательной среде (соли молочной, пировиноградной и янтарной кислот) значительно влияет на продуцирование пропионовой и уксусной кислот культурами P. shermanii. Отношение пропионовой кислоты к уксусной изменяется, с лактатом оно равно 1,83 (17,55:9,55), с пируватом – 0,64 (8,20:12,75) и янтарной кислотой – 2,0 (3,30:1,50). В присутствии лактозы продуцирование пропионовой кислоты происходит более энергично, чем в присутствии глюкозы. При сбраживании культурами P.jensenii пировиноградной кислоты более окисленной, чем молочная, также наблюдается, что соотношение пропионовой и уксусной кислот смещается в сторону более окисленной уксусной. Соотношение кислот зависит от состава закваски. Так, при использовании закваски с двумя культурами – L. helveticum и Str. thermophilus это соотношение ниже, чем при использовании одной L.helveticum.
Таким образом, уникальность пропионовокислого брожения обусловлена: 1) участием ФЕП – карбоксилтрансфосфорилазы фермента, не обнаруженного у других организмов, синтезирующих пропионат; благодаря наличию этого фермента, брожение, осуществляемое пропионово-кислыми бактериями, работает как циклический процесс.
2) особым способом образования пропионата, которое сопряжено с восстановлением фумарата до сукцината и окислением пирувата до ацетата и СО2; транспорт электронов, сопровождающий эти реакции, сопряжен с окислительным фосфорилированием и синтезом АТФ;
3) высокий выход АТФ, превышающий выход АТФ в других известных брожениях. 1,5 М глюкозы могут дать пропионовокислым бактериям около 6 М АТФ.
Энергетическая эффективность пропионовокислого брожения связана также с выработкой пропионовыми бактериями новых метаболических способностей: реакций транскарбоксилирования и перегруппировки, участия в процессе КоА-производных. Образование дикарбоновой кислоты из пировиноградной с использованием механизма транскарбоксилирования вместо прямого карбоксилирования пирувата позволяет избежать дополнительных энергетических затрат на этом этапе брожения. Все это вместе взятое позволяет рассматривать пропионовокислое брожение как более совершенный из рассмотренных до сих пор способов получения энергии в анаэробных условиях.
2. Использование пропионовокислых бактерий в промышленности
Сыроделие – наиболее древняя биотехнология, использующая биохимическую активность пропионовых бактерий. Первые исследования пропионовокислых бактерий были связаны с изучением их роли в созревании сыров. Наиболее высокими органолептическими свойствами и длительными сроками хранения обладают твердые сычужные сыры с высокой температурой второго нагревания, при изготовлении которых принимают участие пропионовокислые бактерии. Общее правило, касающееся использования этих бактерий в созревании сыров, гласит: вреден как недостаток, так и избыток пропионовокислых бактерий, но без их участия сыр нужного качества изготовить невозможно; могут получаться «слепые», т.е. сыры без «глазков» или с другими дефектами. Многие пороки лучших сыров вызваны отсутствием или слабым ростом пропионовокислых бактерий.
Основная роль пропионовокислых бактерий в созревании сыров состоит в использовании лактатов, образованных молочнокислыми бактериями при сбраживании лактозы молока, при этом лактаты превращаются в пропионовую, уксусную кислоты и СО2. Кислоты обеспечивают острый вкус сыров и участвуют в консервации молочного белка казеина; гидролитическое расщепление липидов с образованием жирных кислот важно для развития органолептических свойств сыра; образование пролина и других аминокислот, а также летучих веществ: ацетоина, диацетила, диметилсуль-фида, ацетальдегида, участвующих в формировании аромата сыра; образование углекислоты в процессе пропионово-кислого брожения лактата и декарбоксилирования аминокислот (главным образом); СО2 участвует в создании рисунка сыра - «глазков», образовании витаминов и в первую очередь витамина В12.
Созревание сыра – сложный биохимический процесс, протекающий при участии сычужного фермента, ферментов молока, молочнокислых и пропионовых бактерий. Происходят энзиматические изменения в белках, жире, аминокислотах; формируется аромат, внешний вид, консистенция сыра.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики