Характеристика общих свойств микроорганизмов
Рис. 28. Регуляторные воздействия на уровень клеточных метаболитов (продуктов)
Факторы, регулирующие активность ферментов, разнообразны по своей природе (рис. 28). Физические факторы (температура, давление, свет, магнитное поле, электрические импульсы оказывают менее специфическое действие, чем химические. В свою очередь действие последних также может быть разделено на несколько типов. Одни
химические вещества связываются с активным центром фермента, например субстраты, кофакторы, конкурентные ингибиторы, что приводит к изменению ферментативной активности. Другие вещества взаимодействуют со специальными участками на поверхности молекулы определенного типа фермента, не имеющими непосредственного отношения к центрам каталитической активности, но тем не менее приводящими к ее изменению.
Наконец, активность некоторых ферментов регулируется путем химической модификации их молекулы, в основе которой лежит ковалентное обратимое связывание с ферментом определенной группировки, что приводит к изменению его активности. У прокариот известны две ферментные системы, активность которых регулируется таким путем. Глутаминсинтетаза E. coli, катализирующая синтез глутамина, существует в двух формах, различающихся присутствием в одной из них остатка адениловой кислоты. Присоединение его с помощью ковалентной связи, катализируемое соответствующим модифицирующим ферментом, приводит к образованию менее активной аденилированной глутаминсинтетазы:
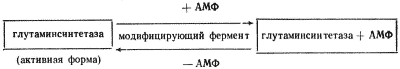
Удаление адениловой группы, ведущее к возникновению деаденилированной формы фермента, резко повышает его каталитическую активность. Аналогичный механизм регулирования активности фермента путем присоединения и удаления остатка уксусной кислоты (ацетилирование — деацетилирование) обнаружен для цитратлиазы у фотосинтезирующей бактерии Rhodopseudomonas gelatinosa. В этом случае активна ацетилированная форма фермента.
Наиболее быстрым, точным и тонким механизмом регуляции активности ферментов является регуляция, которой подвергается определенный тип ферментов, получивших название аллостерических21. Эти ферменты, как правило, занимают ключевые позиции в обмене веществ, располагаясь в "стратегических" пунктах клеточного метаболизма — начале метаболических путей или местах разветвлений, где расходятся или сходятся несколько путей.
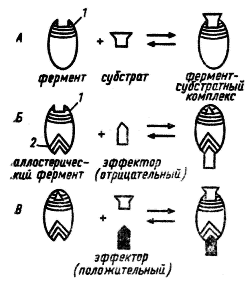
Рис. 29. Связывание субстрата с ферментом (А) и действие отрицательного (Б) и положительного (И) эффектора на каталитическую активность аллостерического фермента: 1 — каталитический центр; 2 — регуляторный центр (no Schlegel, 1972)
Аллостерические ферменты имеют каталитический и регуляторный (аллостерический) центры, пространственно разобщенные, но функционально тесно взаимосвязанные. Каталитическая активность фермента меняется в результате связывания с его регуляторным центром определенных метаболитов, называемых эффекторами. Кроме конечных продуктов данного пути, эффекторами могут быть субстраты ферментов, а также некоторые конечные продукты родственных метаболических путей. Если действие эффектора приводит к понижению каталитической активности фермента, такой эффектор называется отрицательным, или ингибитором. Положительным называют эффектор, действие которого повышает каталитическую активность фермента. Положительным эффектором, или активатором, чаще всего бывает субстрат данного фермента.
Связывание эффектора с регуляторным центром приводит к изменению сродства фермента к субстрату в результате какого-то конформационного изменения фермента (рис. 29).
Наиболее простой случай аллостерической регуляции — регуляция первого фермента неразветвленного биосинтетического пути его конечным продуктом. Если конечный продукт накапливается в избытке, он подавляет активность первого фермента в процессе, называемом ингибированием по принципу обратной связи Примером такого типа регулирования является ингибирование биосинтеза L-изолейцина (рис. 30) Первый фермент на пути синтеза L-изолейцина L-треониндезаминаза является аллостерическим и ингибируется только L-изолейцином.

Рис. 30. Регуляция биосинтеза L-изолейцина по механизму отрицательной обратной связи; ф, — треониндезаминаза; ф — фз — ферменты, катализирующие промежуточные стадии биосинтеза L-изолейцина. Стрелкой показано ингибирование треониндезамнназы L-изолейцином
13. Механизм поглощения субстратов у микроорганизмов.
Основной источник энергии в клетке - окисление субстратов кислородом воздуха. Этот процесс осуществляется тремя путями: присоединением кислорода к атому углерода, отщеплением водорода или потерей электрона. В клетках окисление протекает в форме последовательного переноса водорода и электронов от субстрата к кислороду. Кислород играет в этом случае роль восстанавливающегося соединения (окислителя). Окислительные реакции протекают с высвобождением энергии. Для биологических реакций характерны сравнительно небольшие изменения энергии. Это достигается за счет дробления процесса окисления на ряд промежуточных стадий, что позволяет запасать ее небольшими порциями в виде макроэргических соединений (АТФ). Восстановление атома кислорода при взаимодействии с парой протонов и электронов приводит к образованию молекулы воды.
Тканевое дыхание
Это процесс потребление клетками тканей организма кислорода, который участвует в биологическом окислении. Такой вид окисления называют аэробным окислением. Если конечным акцептором в цепи переноса водорода выступает не кислород, а другие вещества (например пировиноградная кислота), то такой тип окисления называют анаэробным.
Т.о. биологическое окисление - это дегидрирование субстрата с помощью промежуточных переносчиков водорода и его конечного акцептора.
Дыхательная цепь (ферменты тканевого дыхания) - это переносчики протонов и электронов от окисляемого субстрата на кислород. Окислитель - это соединение, способное принимать электроны. Такая способность количественно характеризуется окислительно-восстановительным потенциалом по отношению к стандартному водородному электроду, рН которого равен 7,0. Чем меньше потенциал соединения, тем сильнее его восстанавливающие свойства и наоборот.
Т. о. любое соединение может отдавать электроны только соединению с более высоким окислительно-восстановительным потенциалом. В дыхательной цепи каждое последующее звено имеет более высокий потенциал, чем предыдущее.
Дыхательная цепь состоит из:
1.НАД - зависимой дегидрогеназы;
2.ФАД- зависимой дегидрогеназы;
3.Убихинона (КоQ);
4.Цитохрмов b, c, a+a3 .
НАД-зависимые дегидрогеназы. В качестве кофермента содержат НАД и НАДФ. Пиридиновое кольцо никотинамида способно присоединять электроны и протоны водорода.
ФАД и ФМН-зависимые дегидрогеназы содержат в качестве кофермента фосфорный эфир витамина В2 (ФАД).
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики