Характеристика общих свойств микроорганизмов
Фермент трансальдолаза катализирует перенос остатка диоксиацетона (но не свободного диоксиацетона) от седогептулозо-7-фосфата на гли-церальдегид-3-фосфат:
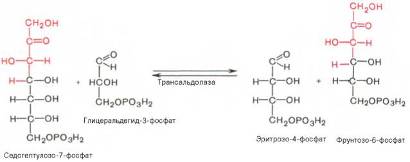
Шесть молекул глюкозо-6-фосфата, вступая в пентозофосфатный цикл, образуют 6 молекул рибулозо-5-фосфата и 6 молекул СО2, после чего из 6 молекул рибулозо-5-фо
сфата снова регенерируется 5 молекул глюко-зо-6-фосфата. Однако это не означает, что молекула глюкозо-6-фосфата, вступающая в цикл, полностью окисляется. Все 6 молекул СО2 образуются из С-1-атомов 6 молекул глюкозо-6-фосфата.
Валовое уравнение окислительной и неокислительной стадий пентозофосфатного цикла можно представить в следующем виде:
![]()
или
![]()
Образовавшийся НАДФН используется в цитозоле на восстановительные синтезы и, как правило, не участвует в окислительном фосфорилировании, протекающем в митохондриях.
15. Особенности пути Ембдена-Мейергофа-Парнаса (гликолиз)
Гексозодифосфатний шлях (гліколіз), або схема Ембдена – Мейєргофа-Парнаса, починається із глюкози і через ряд послідовних реакцій деградації призводить до утворення пірувату . Перша реакція відбувається з витратою енергії у вигляді макроергічного зв'язку АТФ і пов'язана з утворенням фосфорильованого похідного глюкози - глюкозо-6-фосфату. У наступній реакції відбувається ізомеризація глюкозо-6-фосфату у фруктозо-6-фосфат. При дії фосфофруктокінази відбувається утворення фруктозо-1,6-дифосфату. У наступній реакції під впливом альдолази утворюється дві триози: 3-фосфогліцериновий альдегід і фосфодиоксиацетон, які є ізомерами і тому можуть переходити один в одного. Альдолаза фруктозо-1,6-дифосфату є ключовим ферментом для цього шляху, виявлення активності якого чітко вказує на наявність гексозодифосфатного шляху. Наступні реакції пов'язані з окисненням 3-фосфогліцеринового альдегіду під впливом гліцераль-дегід-3-фосфатдегідрогенази з утворенням макроергічного зв'язку 1,3-дифосфо-гліцеринової кислоти. Наявний в цій речовині макроергічний зв'язок перетворюється в макроергічний зв'язок АТФ під час першого субстратного фосфо-рилювання. Друге фосфорилювання пов'язане з перетворенням макроергіч-ної сполуки - фосфоенолпіровиноградної кислоти в піруват. Більш докладно механізми утворення АТФ під час двох субстратних фосфорилювань будуть описані в наступних розділах. Подальша доля пірувату у випадку аеробних процесів пов'язана з перетворенням в ацетил-КоА; в анаеробних умовах від пірувату можуть починатись перетворення за різними схемами процесів бродіння.
Як найбільш показовий приклад можна навести гомоферментативне молочнокисле бродіння, за якого з пірувату утворюється молочна кислота.
Процес спиртового бродіння відбувається за цією ж схемою, але піруват піддається декарбоксилюванню з утворенням ацетоальдегіду, який потім 'Дновлюється НАД+-залежною алкогольдегідрогеназою до етанолу.
Деякі види ентеробактерій здійснюють бродіння з утворенням цілої серії кислот, серед яких домінує мурашина, яка потім може піддаватись утилізації під дією ферменту форміатгідрогенази до С02 і Н2. Такі реакції називають mурашинокислим бродінням. Для ентеробактерій також характерний особли-ним Шлях синтезу етанолу з пірувату через утворення ацетил-КоА з наступним відновленням його до етанолу.
Бродіння з утворенням ацетату може відбуватись декількома шляхам Один з них пов'язаний з відновленням СОг до оцтової кислоти; під час інш го відбувається утворення ацетату з ацетил-КоА.
Більш складним процесом є пропіоновокисле бродіння, яке складається фрагменту циклу Кребса на ділянці від щавлевооцтової до янтарної кислоти Кінцевий продукт бродіння - пропіонова кислота.
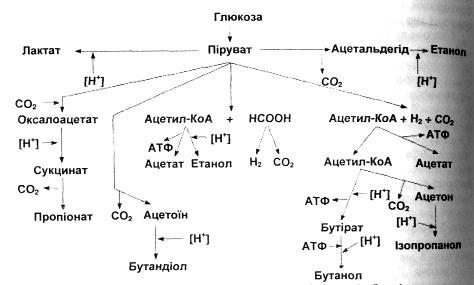
Найбільший спектр кінцевих продуктів утворюється в результаті маслянокислого бродіння: ізопропанол, масляна кислота, бутанол, ацетон, етанол, оцтова кислота.
16. ПУТЬ ЭНТНЕРА — ДУДОРОВА
Общая схема третьего пути расщепления углеводов эубактериями представлена на рис. 67.
Первые два его этапа — фосфорилирование молекулы глюкозы и ее дегидрирование до 6-фосфоглюконовой кислоты — идентичны первым двум этапам окислительного пентозофосфатного пути. Специфичны для пути Энтнера — Дудорова две следующие реакции: 1) дегидратирование 6-фосфоглюконовой кислоты, приводящее к образованию КДФГ-кислоты; 2) расщепление продукта первой реакции на два C3-фрагмента. Конечными продуктами второй реакции являются пировиноградная кислота и 3-ФГА. Последний окисляется в пировиноградную кислоту так же, как в гликолитическом пути. Следовательно, при разложении молекулы глюкозы до пирувата по пути Энтнера — Дудорова образуется 1 молекула АТФ (2 молекулы АТФ синтезируются на отрезке пути 3-ФГА ® пировиноградная кислота минус 1 молекула АТФ, затраченная на фосфорилирование глюкозы), 1 молекула НАД-H2 и 1 молекула НАДФ-H2.
Путь Энтнера — Дудорова имеет важное значение, когда сбраживаемыми субстратами служат глюконовая, маннановая, гексуроновые кислоты или их производные. Он функционирует у довольно широкого круга эубактерии, главным образом, грамотрицательных, получающих энергию в процессе дыхания (энтеробактерии50, виды Azotobacter, Pseudomonas, Alcaligenes, Rhizobium, Spirillum, Xanthomonas, Thiobacillus и др.). У анаэробов он встречается довольно редко. В качестве примера организма, сбраживающего сахара по пути Энтнера — Дудорова, можно привести облигатно анаэробную бактерию Zymomonas mobilis. Однако ее изучение позволяет предполагать, что Z. mobilis — вторичный анаэроб, произошедший от цитохромсодержащих аэробов. Путь Энтнера — Дудорова обнаружен у некоторых клостридиев, что еще раз подчеркивает неоднородность эубактерий, объединенных в эту таксономическую группу.
У энтеробактерий гликолитический и окислительный пентозофосфатный пути функционируют как центральные конститутивные пути метаболизирования углеводов, путь Энтнера — Дудорова — как индуцибельный.
Согласно существующим представлениям путь Энтнера — Дудорова сформировался позднее гликолитического и окислительного пентозофосфатного путей и возник как ответвление последнего, поскольку начала окислительного пентозофосфатного пути и пути Энтнера — Дудорова идентичны и для последнего необходимо было сформировать только два новых фермента (6-фосфоглюконатдегидратазу и КДФГ-альдолазу). Появление пути Энтнера — Дудорова, вероятно, было вызвано высокой потребностью клеток в пирувате, поэтому возникла необходимость сформировать механизм, при помощи которого пируват образовывался бы из исходного субстрата как можно более коротким и прямым путем. Действительно, к получению пирувата по пути Энтнера — Дудорова ведут всего 4 реакции, в то время как в гликолитическом пути для этого требуется 9 ферментативных преобразований.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики