Наследуемый биосинтез белков
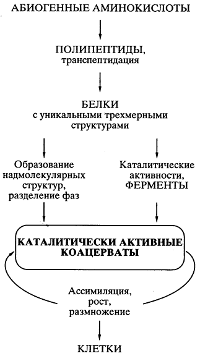
Рис. 5. Схематическое представление пути происхождения жизни согласно белково-коацерватной теории А.И. Опарина
Все было хорошо продумано и научно обосновано в теории, кроме одной проблемы, на которую долго закрывали глаза почти все специалисты в области происхождения жизни. Если спонтанно, путем случайных
безматричных синтезов в коацервате возникали единичные удачные конструкции белковых молекул (например, эффективные катализаторы, обеспечивающие преимущество данному коацервату в росте и размножении), то как они могли копироваться для распространения внутри коацервата, а тем более для передачи коацерватам-потомкам? Теория оказалась неспособной предложить решение проблемы точного воспроизведения - внутри коацервата и в поколениях - единичных, случайно появившихся эффективных белковых структур.
Мир РНК как предшественник современной жизни. Накопление знаний о генетическом коде, нуклеиновых кислотах и биосинтезе белков привело к утверждению принципиально новой идеи о ТОМ, что все начиналось вовсе не с белков, а с РНК [13-15]. Нуклеиновые кислоты являются единственным типом биологических полимеров, макромолекулярная структура которых, благодаря принципу комплементарности при синтезе новых цепей (подробнее см. [3]), обеспечивает возможность копирования собственной линейной последовательности мономерных звеньев, другими словами, возможность воспроизведения (репликации) полимера, его микроструктуры. Поэтому только нуклеиновые кислоты, но не белки, могут быть генетическим материалом, то есть воспроизводимыми молекулами, повторяющими свою специфическую микроструктуру в поколениях.
По ряду соображений именно РНК, а не ДНК, могла представлять собой первичный генетический материал.
Во-первых, и в химическом синтезе, и в биохимических реакциях рибонуклеотиды предшествуют дезоксирибонуклеотидам; дезоксирибонуклеотиды - продукты модификации рибонуклеотидов (см. рис. 2).
Во-вторых, в самых древних, универсальных процессах жизненного метаболизма широко представлены именно рибонуклеотиды, а не дезоксирибонуклеотиды, включая основные энергетические носители типа рибонуклеозид-полифосфатов (АТФ и т.п.).
В-третьих, репликация РНК может происходить без какого бы то ни было участия ДНК, а механизм редупликации ДНК даже в современном живом мире требует обязательного участия РНК-затравки в инициации синтеза цепи ДНК.
В-четвертых, обладая всеми теми же матричными и генетическими функциями, что и ДНК, РНК способна также к выполнению ряда функций, присущих белкам, включая катализ химических реакций. Таким образом, имеются все основания рассматривать ДНК как более позднее эволюционное приобретение - как модификацию РНК, специализированную для выполнения функции воспроизведения и хранения уникальных копий генов в составе клеточного генома без непосредственного участия в биосинтезе белков.
После того как были открыты каталитически активные РНК, идея первичности РНК в происхождении жизни получила сильнейший толчок к развитию, и была сформулирована концепция самодостаточного мира РНК, предшествовавшего современной жизни [16, 17]. Возможная схема возникновения мира РНК представлена на рис. 6.
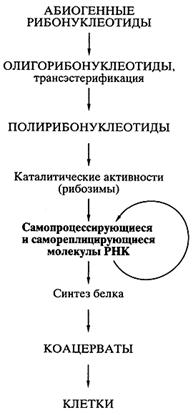
Рис. 6. Схематическое представление пути происхождения жизни согласно современной концепции первичности мира РНК
Абиогенный синтез рибонуклеотидов и их ковалентное объединение в олигомеры и полимеры типа РНК могли происходить приблизительно в тех же условиях и в той же химической обстановке, что постулировались для образования аминокислот и полипептидов. Недавно А.Б. Четверин с сотрудниками (Институт белка РАН) экспериментально показали, что по крайней мере некоторые полирибонуклеотиды (РНК) в обычной водной среде способны к спонтанной рекомбинации, то есть обмену отрезками цепи, путем транс-эстерификации [18]. Обмен коротких отрезков цепи на длинные, должен приводить к удлинению полирибонуклеотидов (РНК), а сама подобная рекомбинация способствовать структурному многообразию этих молекул. Среди них могли возникать и каталитически активные молекулы РНК.
Даже крайне редкое появление единичных молекул РНК, которые были способны катализировать полимеризацию рибонуклеотидов или соединение (сплайсинг) олигонуклеотидов на комплементарной цепи как на матрице [19, 20], означало становление механизма репликации РНК. Репликация самих РНК-катализаторов (рибозимов) должна была повлечь за собой возникновение самореплицирующихся популяций РНК. Продуцируя свои копии, РНК размножались. Неизбежные ошибки в копировании (мутации) и рекомбинации в самореплицирующихся популяциях РНК создавали все большее разнообразие этого мира. Таким образом, предполагаемый древний мир РНК - это "самодостаточный биологический мир, в котором молекулы РНК функционировали и как генетический материал, и как энзимоподобные катализаторы" [21].
Возникновение биосинтеза белка. Далее на основе мира РНК должно было происходить становление механизмов биосинтеза белка, появление разнообразных белков с наследуемой структурой и свойствами, компартментализация систем биосинтеза белка и белковых наборов, возможно, в форме коацерватов и эволюция последних в клеточные структуры - живые клетки (см. рис. 6).
Проблема перехода от древнего мира РНК к современному белок-синтезирующему миру - наиболее трудная даже для чисто теоретического решения. Возможность абиогенного синтеза по-липептидов и белковоподобных веществ не помогает в решении проблемы, так как не просматривается никакого конкретного пути, как этот синтез мог бы быть сопряжен с РНК и подпасть под генетический контроль. Генетически контролируемый синтез полипептидов и белков должен был развиваться независимо от первичного абиогенного синтеза, своим путем, на базе уже существовавшего мира РНК. В литературе предложено несколько гипотез происхождения современного механизма биосинтеза белка в мире РНК, но, пожалуй, ни одна из них не может рассматриваться как детально продуманная и безупречная с точки зрения физико-химических возможностей. Представлю свою версию процесса эволюции и специализации РНК, ведущего к возникновению аппарата биосинтеза белка (рис. 7), но и она не претендует на законченность.
Предлагаемая гипотетическая схема содержит два существенных момента, кажущихся принципиальными.
Во-первых, постулируется, что абиогенно синтезируемые олигорибонуклеотиды активно рекомбинировали посредством механизма спонтанной неэнзиматической трансэстерификации [18], приводя к образованию удлиненных цепей РНК и давая начало их многообразию. Именно этим путем в популяции олигонуклеотидов и полинуклеотидов и могли появиться как каталитически активные виды РНК (рибозимы), так и другие виды РНК со специализированными функциями (см. рис. 7). Более того, неэнзиматическая рекомбинация олигонуклеотидов, комплементарно связывающихся с полинуклеотидной матрицей, могла обеспечить сшивание (сплайсинг) фрагментов, комплементарных этой матрице, в единую цепь. Именно таким способом, а не катализируемой полимеризацией мононуклеотидов, могло осуществляться первичные копирование (размножение) РНК. Разумеется, если появлялись рибозимы, обладавшие полимеразной активностью [20], то эффективность (точность, скорость и продуктивность) копирования на комплементарной. матрице должна была значительно возрастать.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики