Наследуемый биосинтез белков
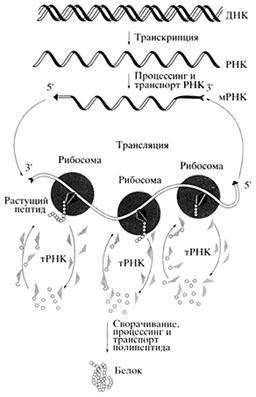
Рис. 1. Общая схема биосинтеза белков
Однако, чтобы синтезировать белки, одной только информации или программы недостаточно - нужен еще и материал, из которого их можно делать. Поток материала для синтеза белков идет в рибосомы через посредство третьего класса клеточных РНК - РНК-переносчиков (trans
fer RNA, транспортные РНК, тРНК). Они ковалентно связывают - акцептируют - аминокислоты, которые служат строительным материалом для беЛков, и в виде аминоацил-тРНК поступают в рибосомы. В рибосомах аминоацил-тРНК взаимодействуют с кодонами - трехнуклеотидными комбинациями - мРНК, в результате чего и происходит декодирование кодонов в процессе трансляции.
Рибонуклеиновые кислоты
Итак, перед нами набор главных клеточных РНК, определяющих основной процесс современной живой материи - биосинтез белка. Это мРНК, рибосомные РНК и тРНК. РНК синтезируются на ДНК с помощью ферментов - РНК-полимераз, осуществляющих транскрипцию - переписывание определенных участков (линейных отрезков) двутяжевой ДНК в форму однотяжевой РНК. Участки ДНК, кодирующие клеточные белки, переписываются в виде мРНК, тогда как для синтеза многочисленных копий рибосомной РНК и тРНК имеются специальные участки клеточного генома, с которых идет интенсивное переписывание без последующей трансляции в белки.
Химическая структура РНК. Химически РНК очень похожа на ДНК. Оба вещества - это линейные полимеры нуклеотидов. Каждый мономер - нуклеотид - представляет собой фосфорилированный N-гликозид, построенный из остатка пятиуглеродного сахара - пентозы, несущего фосфатную группу на гидроксильной группе пятого углеродного атома (сложноэфирная связь) и азотистое основание при первом углеродном атоме (N-гликозидная связь). Главное химическое различие между ДНК и РНК состоит в том, что сахарный остаток мономера РНК - это рибоза, а мономера ДНК - дезоксирибоза, являющаяся производным рибозы, в котором отсутствует гидроксильная группа при втором углеродном атоме (рис. 2).
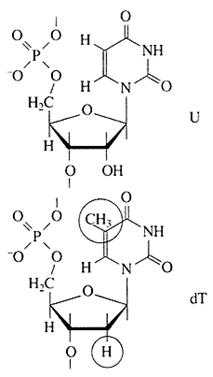
Рис. 2. Химические формулы остатков одного из рибонуклеотидов - уридиловой кислоты (U) и гомологичного ему дезоксирибонуклеотида - тимидиловой кислоты (dT)
Азотистых оснований и в ДНК, и в РНК четыре вида: два пуриновых - аденин (А) и гуанин (G) -и два пиримидиновых - цитозин (С) и урацил (U) или его метилированное производное тимин (Т).
Урацил характерен для мономеров РНК, а тимин - для мономеров ДНК, и это второе различие РНК и ДНК. Мономеры - рибонуклеотиды РНК или дезоксирибонуклеотиды ДНК - образуют полимерную цепь посредством формирования фосфодиэфирных мостиков между сахарными остатками (между пятым и третьим атомами углерода пентозы). Таким образом, полимерная цепь нуклеиновой кислоты - ДНК или РНК - может быть представлена как линейный сахаро-фосфатный остов с азотистыми основаниями в качестве боковых групп.
Макромолекулярная структура РНК. Принципиальное макроструктурное различие двух типов нуклеиновых кислот состоит в том, что ДНК - это единая двойная спираль, то есть макромолекула из двух комплементарно связанных полимерных тяжей, спирально закрученных вокруг общей оси (см. [1, 3]), а РНК - однотяжевой полимер. В то же время взаимодействия боковых групп - азотистых оснований - друг с другом, а также с фосфатами и гидроксилами сахаро-фосфатного остова приводят к тому, что однотяжевой полимер РНК сворачивается на себя и скручивается в компактную структуру [4], подобно сворачиванию полипептидной цепи белка в компактную глобулу. Таким способом уникальные нуклеотидные последовательности РНК могут формировать уникальные пространственные структуры.
Впервые специфическая пространственная структура РНК была продемонстрирована при расшифровке атомной структуры одной из тРНК в 1974 г. [5,6] (рис. 3). Сворачивание полимерной цепи тРНК, состоящей из 76 нуклеотидных мономеров, приводит к формированию очень компактного глобулярного ядра, из которого под прямым углом торчат два выступа. Они представляют собой короткие двойные спирали по типу ДНК, но организованные за счет взаимодействия участков одной и той же цепи РНК. Один из выступов является акцептором аминокислоты и участвует в синтезе полипептидной цепи белка на рибосоме, а другой предназначен для комплементарного взаимодействия с кодирующим триплетом (кодоном) мРНК в той же рибосоме. Только такая структура способна специфически взаимодействовать с белком-ферментом, навешивающим аминокислоту на тРНК, и с рибосомой в процессе трансляции, то есть специфически "узнаваться" ими.

Рис. 3. Атомная (слева) и скелетная (справа) модели фенилаланиновой тРНК дрожжей
Изучение изолированных рибосомных РНК дало следующий разительный пример формирования компактных специфических структур из еще более длинных линейных полимеров этого типа. Рибосома состоит из двух неравных частей - большой и малой рибосомных субчастиц (субъединиц). Каждая субчастица построена из одной высокополимерной РНК и целого ряда разнообразных рибосомных белков. Длина цепей рибосомных РНК весьма значительна: так, РНК малой субчастицы бактериальной рибосомы содержит более 1500 нуклеотидов, а РНК большой субчастицы - около 3000 нуклеотидов. У млекопитающих, включая человека, эти РНК еще больше - около 1900 нуклеотидов и более 5000 нуклеотидов в малой и большой субчастицах соответственно.
Было показано, что изолированные рибосомные РНК, отделенные от их белковых партнеров и полученные в чистом виде, сами способны спонтанно сворачиваться в компактные структуры, по своим размерам и форме похожие на рибосомные субчастицы [7]. Форма большой и малой субчастиц разная, и соответственно различается форма большой и малой рибосомных РНК (рис. 4). Таким образом, линейные цепи рибосомной РНК самоорганизуются в специфические пространственные структуры, определяющие размеры, форму и, по-видимому, внутреннее устройство рибосомных субчастиц, а следовательно, и всей рибосомы.
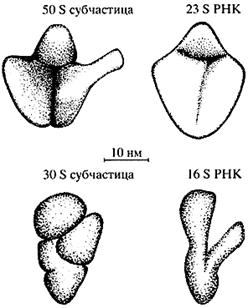
Рис. 4. Сравнение контуров рибосомных субчастиц бактерий и их изолированных высокополимерных РНК в компактной форме по данным электронной микроскопии: вверху - большая субчастица и ее РНК; внизу - малая субчастица и ее РНК
Минорные РНК. По мере изучения компонентов живой клетки и отдельных фракций тотальной клеточной РНК выяснялось, что тремя главными видами РНК дело не ограничивается. Оказалось, что в природе существует множество других видов РНК. Это, в первую очередь, так называемые "малые РНК", которые содержат до 300 нуклеотидов, часто с неизвестными функциями. Как правило, они ассоциированы с одним или несколькими белками и представлены в клетке в виде рибонуклеопротеидов - "малых РНП" [8].
Малые РНК присутствуют во всех отделах клетки, включая цитоплазму, ядро, ядрышко, ми-тохондрии. Большая часть тех малых РНП, функции которых известны, участвует в механизмах посттранскрипционной обработки главных видов РНК (RNA processing) - превращении предшественников мРНК в зрелые мРНК (сплайсинг), редактировании мРНК, биогенезе тРНК, созревании рибосомных РНК. Один из наиболее богато представленных в клетках видов малых РНП (SRP) играет ключевую роль в транспорте синтезируемых белков через клеточную мембрану. Известны виды малых РНК, выполняющих регуляторные функции в трансляции. Специальная малая РНК входит в состав важнейшего фермента, ответственного за поддержание редупликации ДНК в поколениях клеток - теломеразы. Следует сказать, что их молекулярные размеры сопоставимы с размерами клеточных глобулярных белков. Таким образом, постепенно становится ясно, что функционирование живой клетки определяется не только многообразием синтезируемых в ней белков, но и присутствием богатого набора разнообразных РНК, из которых малые РНК в значительной мере имитируют компактность и размеры белков.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики