Стратегии изучения зрительной коры
Каким образом сигналы, определяющие маленькие, яркие, темные или цветовые пятна на сетчатке могут быть преобразованы в сигналы, которые несут информацию о форме, размере, цвете, движении и глубине расположения объектов. Методики, считающиеся сейчас рутинными, такие как оптическая регистрация, введение пероксидазы хрена или сканирование мозга, в ту пору даже не могли быть представлены. В начал
е работы перед Хьюбелем и Визелем стояли совершенно неизученные вопросы, которые исходили из их предположения, что зрительные центры в коре осуществляют переработку информации подобно тому, как это происходит в сетчатке, только на более высоком уровне.
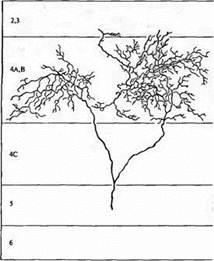
Рис. 1. Аксон "off" центра из ЛКТ, заканчивающийся в слое 4 зрительной коры кошки. В аксон при помощи микроэлектрода была введена пероксидаза хрена. Терминали сгруппированы в два кластера, разделенных неокрашенной зоной, получающей иннервацию от другого глаза.
Одной из ключевых стратегий в их анализе было использование стимулов, которые имитировали сигналы, возникающие в нормальных условиях. Например, края, контуры, а также простые паттерны, представленные глазу, позволяли выявлять особенности организации, которые никогда бы не были обнаружены, используя яркие вспышки света, не имеющие какой-либо определенной формы. Другим ключевым моментом успеха подхода Хьюбеля и Визеля было то, что они искали не просто стимул, которые может вызвать ответ определенного нейрона, а наиболее эффективный для этого стимул. Исследование этой проблемы на разных этапах зрительной системы выявило много удивительных и неожиданных результатов. В ранних работах было показано, что рецептивные поля простых и сложных клеток в первичной зрительной коре составляют начальные этапы распознавания паттернов. Кроме того, анализ рецептивных полей четко выявил полезный и упрощающий принцип, что нейроны, расположенные вдоль радиальных трактов, идущих от поверхности коры в белое вещество, решают сходные задачи, формируя функциональные колонки. Эти результаты были окончательно подтверждены при помощи длинного электрода, вводимого далеко вглубь коры, одновременно с чем проводилась регистрация сигналов и идентификация свойств клеток, в зависимости от типа возбуждающего их стимула. После регистрации проводилась тщательная гистологическая реконструкция путей прохождения электрода.
Рецептивные поля коры
Сигналы кортикальных нейронов, подобно сигналам от ганглиозных клеток сетчатки и клеток ЛКТ, в основном появляются на фоне постоянной активности клеток. Наблюдения показывают, что диффузное освещение сетчатки слабо влияет на разряды кортикальных нейронов. Почти полная нечувствительность их к диффузному освещению является результатом процесса, отмеченного еще в сетчатке и в ЛКТ. Это результат в равной степени выраженного антагонистического действия тормозных и возбуждающих регионов рецептивных полей кортикальных клеток. Степень активности нейрона изменяется только при соблюдении определенных условий относительно расположения и формы стимула на сетчатке. Рецептивные поля большинства кортикальных нейронов имеют конфигурацию, отличную от таковой у клеток сетчатки и нейронов ЛКТ, поэтому отдельные пятна света часто являются слабыми стимулами (или вообще ими не являются). В своей нобелевской речи, Хьюбель описал эксперимент, благодаря которому они с Визелем впервые сумели заметить это важнейшее свойство2
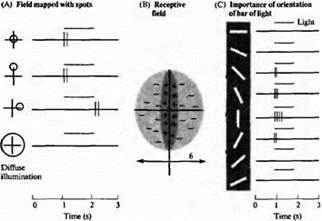
Рис. 2. Ответы простых клеток полосатой коры на пятна (А) и полоски (С) света. Рецептивные поля (В) имеют узкую центральную "оп"-зону (+), окруженную симметрично антагонистическими "off''-зонами (-). Наилучшим стимулом для данной клетки является вертикально ориентированная полоска света в центре рецептивного поля (пятая запись сверху на С). Другие ориентации менее эффективны или вовсе неэффективны. Рассеянный свет не является каким-либо стимулом. Полоска над каждой записью в А и С указывает продолжительность стимула.
Следуя серии маленьких удачных находок, Хьюбель и Визель нашли наиболее подходящие стимулы света для различных клеток коры; первоначально они классифицировали рецептивные поля на простые и сложные. Каждая из этих категорий включала большое количество подгрупп и важных переменных, которые определяли механизмы восприятия.
Ответы простых клеток
Большинство простых клеток расположено в слоях 4 и 6, а также в глубине слоя 3. Все эти слои получают сигналы непосредственно из ЛКТ (хотя именно слой 4С является наиболее типичным местом, куда направляются отростки из ЛКТ, как мы уже ранее указывали). Рецептивные поля простых клеток могут быть определены при помощи пятен света, проецируемых в определенные области сетчатки, и они имеют несколько разновидностей. Один тип простых клеток имеет рецептивное поле, которое состоит из удлиненной узкой центральной части, окруженной двумя антагонистическими зонами. Центр может быть либо возбуждающим, либо ингибирующим. На рис. 2 показано рецептивное поле простой клетки в полосатой коре, определенное при помощи пятен света, которые были способны лишь слабо возбудить клетки в центре поля (потому как они покрывали лишь малую часть центральной "on"-зоны).

Рис. 3. Рецептивное поле простой клетки по лосатой коры. В действительности для каждого типа поля были исследованы все возможные ориентации. Наиболее оптимальным стимулом являлось освещение в виде узкой щели или полоски в центре (для А); темная полоска была оптимальной для В и С;
Необходимые условия для активации одной простой клетки соответствуют тем, что проиллюстрированы на рис. 2. Для оптимальной активации ей необходима полоска света, размерами не более центральной зоны, которая полностью ее заполняет по длине и ширине и ориентирована под определенным углом. Освещение окружающей зоны подавляет текущую активность клетки, а также уменьшает эффективность одновременного освещения центра зоны. Как было предсказано картированием при помощи световых пятен, вертикально ориентированная полоска является наиболее эффективным стимулом. Даже небольшое отклонение от этого паттерна приводит к уменьшению сигнала. Различные клетки имеют рецептивные поля, требующие широкого диапазона различных ориентаций и расположения. Таким образом, при повороте стимула или при смещении его в пределах зрительного поля, активируется другая популяция простых клеток. Распределение ингибирующих и возбуждающих сторон у различных простых клеток может и не быть симметричным, а также поле может состоять из двух продольных регионов, расположенных друг напротив друга — одного возбуждающего, а другого ингибирующего.
На рис. 3 приведены примеры четырех подобных рецептивных полей, имеющих общую ось ориентации, но с различным распределением возбуждающих и угнетающих зон в пределах поля. Максимальный ответ от рецептивного поля, показанного на рис. 3 А, вызывает узкий щелевидный пучок, ориентированный на 1 час по воображаемому циферблату (при условии, что все рецептивное поле представляет из себя циферблат, разделенный на 12 частей). Темная полоска в том же самом месте с ярким освещением по периферии подавляет постоянную активность клетки. Клетки, имеющие поле, иллюстрированное на рис. 3В и С, отвечают на темную полосу в центральной зоне. Для поля, показанного на рис. 3, границы между светом слева и тенью справа является наиболее эффективным "on"-ответом, а освещение, наоборот, оптимально для "off"-разрядов. В простых клетках оптимальная ширина узкого пучка света или полоски тени сравнима с диаметрами регионов с "on"- и "off"-центрами для рецептивных полей в форме бублика у ганглиозных клеток и клеток ЛКТ. Таким образом, кортикальные клетки, которые имеют рецептивные поля в области зрительного пятна, лучше всего возбуждаются узкими полосками света, по сравнению с клетками, имеющими рецептивные поля на периферии сетчатки, что обусловлено более маленькими рецептивными полями ганглиозных клеток ямки.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики