Противостояние организма изменениям барических и термических условий
Приспособление к гипоксии
Мы уже знаем, что экстренное приспособление к высотной гипоксии вызывает прежде всего ряд физиологических реакций, направленных на возможно более полное удовлетворение потребности организма в кислороде. Но этого недостаточно даже на сравнительно небольших высотах: уже на уровне 3500 м парциальное давление кислорода в альвеолах легких снижается на 53%, а насыщение
им крови – на 20%.
Разные органы и составляющие их клетки в неодинаковой степени чувствительны к недостатку кислорода. Особенно страдает от этого центральная нервная система, в частности нервные клетки ее высшего отдела – коры головного мозга. На небольших высотах человек этого еще не замечает, но инструментальные физиологические исследования показывают, что уже на высоте 1500 м у человека ухудшается темновая адаптация, т.е. приспособление глаза к ясному зрению при переходе от яркого освещения к полутемноте. На высотах 2000–2200 м отмечаются нарушения и в функциях коры головного мозга: увеличивается скрытый период речевых реакций, а сама речь становится более примитивной, нарушается тонкая дифференцировка восприятий. На высоте 4000 м хуже протекают условные рефлексы, а выше 6000 м в коре мозга возникает разлитое торможение, появляется сонливость и, наконец, теряется сознание. Но ведь на высоте надо и двигаться, и нередко совершать значительную работу. А это еще более увеличивает потребность в кислороде.
Общеизвестно, что энергию, необходимую для всех проявлений жизнедеятельности, организм получает путем окисления различных веществ, прежде всего углеводов и жиров, накапливая ее посредством окислительного фосфоридирования в легкоутилизируемой форме АТФ. Процесс окисления заключается в отнятии водорода от окисляемых веществ и перенесении его протона и электрона через дыхательную цепь на кислород. Перенос каждой пары протонов и электронов сопровождается образованием трех молекул АТФ. При этом, чем богаче водородом окисляемое вещество, тем больше выход АТФ: при окислении глюкозы образуется 38 молекул АТФ, а при окислении жирных кислот, например пальмитиновой, – 136. А если кислорода недостаточно? Если протоны и электроны движутся по дыхательной цепи, а отдавать их некому? Если вся дыхательная цепь оказывается восстановленной, т.е. забита электронами и протонами? Тогда на помощь «спешит» гликолиз.
Гликолиз происходит следующим образом. Глюкоза подвергается фосфорилированию. Образовавшийся глю-козо-фосфорный эфир после ряда промежуточных превращений и внутримолекулярных перегруппировок расщепляется на две молекулы фосфотриозы, которые подвергаются окислению. От них отнимается водород, но принимает его не кислород, а кофермент НАД. Конечным же продуктом является ПВК. Дальнейшая судьба этого соединения неодинакова и зависит от степени обеспечения организма кислородом: если его достаточно, то ПВК окисляется до СО2 и H2O, а водород с НАД · H2 через дыхательную цепь переносится на кислород; при нехватке кислорода НАД · H2 отдает свой водород ПВК. В результате из ПВК образуется молочная кислота, а НАД способен уже принимать новые порции водорода, поддерживая тем гликолиз: СН3-СО-СООН + НАД. H2-^CH3-СНОН-СООН + НАД.
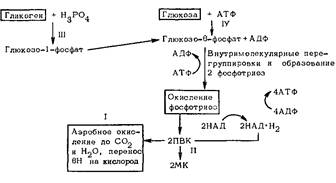
Схема 8. Аэробное и анаэробное окисление глюкозы
MK – молочная кислота. // – аэробное окисление; // – гликолиз; 77/ – путь от гликогена; IV – путь от глюкозы
На первых порах приспособления к гипоксии интенсивность гликолиза возрастает, увеличивается синтез его ферментов. Повышенными возможности гликолиза остаются и при длительном приспособлении к гипоксии. Но у гликолиза три существенных недостатка. Во-первых, аэробно окисляться могут самые различные вещества, а гликолизируется только глюкоза; запасы же гликогена в организме ограниченны. Во-вторых, гликолиз энергетически малоэффективен. В-третьих, организму прежде всего кровь, наводняется молочной кислотой; положение ухудшается еще и тем, что в результате неполного окисления жирных кислот из них образуется в-оксимасляная и ацетоуксусная кислоты, сдвигающие, как и молочная кислота, реакцию внутренней среды в кислую сторону и способные превращаться в ядовитый для организма ацетон.
Естественно, что сдвиг реакции среды в кислую сторону для организма неблагоприятен: если он велик, многие ферменты будут работать не в условиях оптимума кислотности, а значит, проявлять кемаксимальную активность: внутри молекул многих белков могут нарушиться водородные и ионные связи, в результате чего произойдет снижение или даже утрата функциональных свойств данных белков. Конечно, у организма есть средства противостоять этому. Кровь и ткани обладают резервной щелочностью, которая может регулировать реакцию их среды, что осуществляется буферными системами, которые способны связывать при изменении реакции в кислую сторону и OH – при сдвиге в щелочную. Примером может служить бикарбонатнэя буферная система: при избытке H+ бикарбонат, присоединяя их, переходит в плохо диссоциирующую угольную кислоту, а при избытке ОН» угольная кислота превращается в ее двууглекислую соль, а ее водород, соединяясь с гидроксидом, образует крайне слабо диссоциирующую воду. Таким же действием обладают и другие буферные системы: фосфатная, состоящая из одно- и двузамещенной солей фосфорной кислоты, и белки плазмы крови, поскольку они имеют кислую группу – карбоксил и щелочную аминогруппу. Первая связывает ОН-, образуя воду, и переходит в COO-, а вторая – H+, прекращаясь в NH3». Буферным веществом является также и гемоглобин. На долю бикарбонатов приходится 53% всей буферной емкости крови, на долю фосфатов – 5%, белков – 7, гемоглобина – 35%. Таким образом, буферные вещества компенсируют сдвиги реакции среды в кислую или щелочную сторону, убирая, связывая ионы H и ОН.
Так как на высотах понижено парциальное давление не только кислорода, но и углекислоты, организм при усиленном дыхании теряет ее в повышенных количествах. Это тоже уменьшает кислотность крови и может привести даже к сдвигу реакции ее в щелочную сторону. Но потери углекислоты вызывают падение возбудимости дыхательного центра в мозгу и ослабление дыхания. Буферные же системы имеют определенную емкость, и она может быть исчерпана. Реакция снова сдвигается в кислую сторону, что называют некомпенсированным ацидозом.
Для борьбы с избытком молочной кислоты организму остается возможность в малой части выделить ее с мочой, а все остальное – химически устранить, что происходит двумя путями. Первый – окисление молочной кислоты до CO2 и Н2О в печени и мышце сердца, но для этого необходимо повышенное поглощение кислорода, что исключено во время пребывания на высоте и возможно лишь при возвращении на более низкие уровни. Второй путь – гликонеогенез, т.е. переработка молочной кислоты в глюкозу и гликоген. Этот цикл реакций, названный по фамилии открывшего его американского биохимика К. Кори циклом Кори, предоставляет орга-ёнтзму ряд преимуществ. Так, устраняется не только молочная кислота, но и некоторое количество H+, присоединяемое к продуктам превращения молочной кислоты в глюкозу, а также увеличивается снабжение центральной нервной системы необходимой ей глюкозой. Таким образом, цикл Кори – один из важных приспособительных механизмов энергетического обмена.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики