Изучение вопросов биотехнологии в курсе химии средней школы
– А Т Г Ц А А Т Т Ц Т Г А Г А Т Ц Ц А Т А Ц Г
Т А Ц Г Т Т А А Г А Ц Т Ц Т А Г Г Т А Т Г Ц
Фрагмент 1 Линкер Фрагмент 2
Рис. 2. Объединение фрагментов линкерами
Линкерные фрагменты не только обеспечивают объединение генов, но и обусловливают их экспрессию, в связи с чем, часто в середину линкера помещают какой-либо регуляторный генетический элемент, например промотор, или участок
связывания с рибосомой.
После того как рекомбинантная ДНК сшита, ее вводят в живые клетки. При этом рекомбинантные ДНК становятся составной частью генетического аппарата реципиентного организма и, кроме того, они привносят в него новые генетические и физиолого-биохимические свойства, полезные для человека. Но поскольку она не способна к самовоспроизведению, её разрушают внутриклеточные нуклеазы. Для того чтобы рекомбинантная ДНК стала составной частью генетического аппарата клетки, она должна либо встроиться (интегрироваться) в её геном и реплицирваться за его счет, либо быть способной к автономной репликации. Принято молекулы ДНК, способные акцептировать чужеродную ДНК и автономно реплицироваться, называть векторными молекулами. К числу векторов относят плазмиды, бактериофаги, вирусы животных. Векторы должны обладать следующими особенностями:
1. Иметь субстратные участки для определенных эндонуклеаз рестрикции.
2. Иметь свойства репликона.
3. Содержать один или несколько маркерных генов, которые после проникновения вектора в клетку придают ей фенотип, свидетельствующий о присутствии вектора.
Таким образом, все векторы обеспечивают репликацию встроенных генов, их экспрессию, интеграцию в хромосому клетки и т.д. Чаще других в генетической инженерии в качестве векторов используют плазмиды. Плазмидами называют стабильно наследуемые бактериальные репликоны (внехромосомные элементы наследственности). Они представляют собой двуцепочечные кольцевые молекулы ДНК с вариабельными молекулярными массами. Они детерминируют разные свойства: резистентность к антибиотикам (R-плазмиды); биодеградацию (D-плазмиды) и др. Например, плазмиды стафилококков несут гены устойчивости к пенициллину, соединениям ртути и др. Количество плазмид в клетке может колебаться от одной до более ста.
Первый плазмидный вектор был получен С. Коэном (1973). Его источником была плазмида E. coli R6-5 c Mr 65 кДа. Плазмида стала родоначальником серии векторов и других структур.
Плазмида pBR313 содержит уникальные участки расщеплений нескольких рестриктаз (рис. 3).
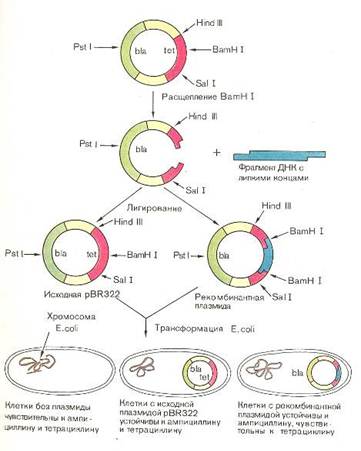
Рис. 3. Схема использования плазмиды pBR322 для отбора клеток, содержащих рекомбинантные плазмиды[14].
Возможности плазмид для генетической инженерии не беспредельны, что связано с их небольшими размерами. Когда нужно клонировать крупные фрагменты ДНК, удобнее использовать векторы на основе бактериофага λ. Геном фага λ досконально изучен[9]. В центральной части линейного генома фага содержится область, необязательная для литической инфекции, которую и используют для вставки клонируемой ДНК. ДНК фага λ разрезают с помощью рестриктазы Eco RI, удаляют необязательную центральную область и на её место встраивают нужный фрагмент ДНК, получая, таким образом, конкамер – предшественник для упаковки фаговой ДНК в зрелые фаговые частицы. Фаг λ очень пластичен: без нарушения развития фага из него можно убрать до 25% ДНК или пристроить до 6% лишней ДНК. В этот фаговый вектор можно встраивать до 23 000 н.п.
В тех случаях, когда не хватает возможностей фага, используют ещё более крупные векторы – космиды. В состав космид входят: ген (маркер) резистентности к антибиотикам, репликон плазмиды и фрагмент ДНК фага λ (так называемый cos-участок). Этот фрагмент представляет собой однонитевые комплементарные участки на концах фаговой ДНК, т.е. «липкие концы».
Ещё один вид векторов – фазмиды, искусственные гибриды между фагом и плазмидой. После встройки чужеродной ДНК они в одних условиях могут размножаться как фаги, а в других – как плазмиды.
Векторы на основе нитевидных фагов применяют тогда, когда удобнее работать с одной цепью ДНК. Так, фага М13 и fd содержат кольцевую одноцепопочечную ДНК с полностью изученными последовательностями нуклеотидов.
Дрожжи Saccharomyces cerevisiae представляют собой перспективные модели для экспериментов с рекомбинантными ДНК. Их гаплоидный геном содержит 1,4 х 107 н.п. (в 3 раза больше, чем у E.coli), распределенных по 17 хромосомам. Дрожжи не инфицируются вирусами, но в их клетках выявлена плазмида, используемая в качестве вектора – 2 мкм-плазмидная ДНК. Важной особенностью дрожжевых систем является то обстоятельство, что клонируемые фрагменты ДНК способны к рекомбинации с гомологичными участками дрожжевого генома, что ведет к стабильной сайт-специфической трансформации последнего (независимо от сохранения или утраты вектора).
Векторные плазмиды и векторные вирусы со встроенными чужеродными генами часто называют гибридными (или химерными) плазмидами (или фагами). После конструирования рекомбинантных ДНК их с помощью трансформации вводят в реципиентный организм: бактериальную, грибную, растительную или животную клетку. Трансформация предусматривает предварительную обработку клеток соединениями(CaCl2), обусловливающими проникновение ДНК внутрь клеток с последующим их помещением в среду, в которой способны существовать только клетки, получившие векторную молекулу, например в среду с определенным антибиотиком.
Процесс инфицирования клеток с помощью чужеродных ДНК, приводящий к образованию зрелого фагового потомства, назван трансфекцией.
Однако эффективность проникновения экзогенной ДНК в клетку довольно низка. Поэтому среди бактерий, подвергшихся трансформации, только небольшая часть оказывается трансформированной. Отделение её от общей массы возможно также в процессе клонирования. Для клонирования бактериальную суспензию определенной концентрации выливают на твердую питательную среду, например на агар с питательными добавками в чашке Петри из расчета 5–10 бактерий на 1 см2 поверхности. Бактериальная клетка на поверхности агара начинает делиться с образованием в итоге маленькой колонии, похожей на шляпку гриба. Эта колония называется клоном, причем из каждой клетки образуется свой клон, все клетки которого имеют свойства бактерии-родоначальника.
Рекомбинантные клоны могут быть идентифицированы по синтезируемому ими продукту (иммунологический метод Брума–Джилберта) [4]. Но чаще приходится идентифицировать непосредственно нуклеотидную вставку с использованием методов гибридизации (Грюнштейн – Хогнесс) [18]. (рис. 4).
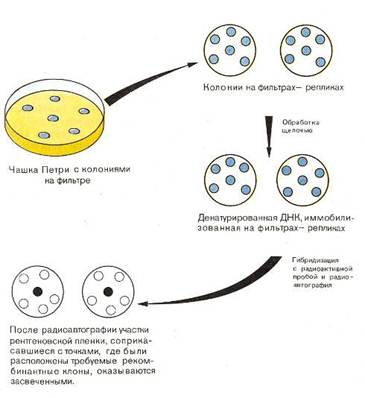
Рис. 4. Метод радиоавтографии в применении к поиску рекомбинантных клонов[14].
Из фрагментов вирусных и бактериальных хромосом уже выделен целый ряд генов. Что же касается выделения специфических генов из фрагментированных эукариотических хромосом, то реализация этой процедуры все еще остается сложной и трудоемкой задачей. Существует два основных подхода для получения специфических генов, подлежащих затем рекомбинации и клонированию. В одном из них, который получил название «шотган» (от англ. shotgun – дробовик), всю клеточную ДНК обрабатывают рестриктирующей эндонуклеазой, образующей в местах разрыва выступающие концы. Полученные фрагменты ДНК встраивают затем в плазмиды E.coli, «раскрытые» (т.е. переведенные в линейную форму) с помощью той же самой рестриктирующей эндонуклеазы. В результате образуется чрезвычайно сложная смесь, состоящая, вероятно, из тысяч разных рекомбинантных плазмид, среди которых лишь одна может содержать нужный ген. Для поиска плазмиды, несущей этот ген, разработаны специальные процедуры, которые называют скринингом.
Другие рефераты на тему «Педагогика»:
- Коррекция нарушений речи у детей с задержкой психического развития
- Выбор профессии. Юриспруденция
- Формирование дивергентного мышления старшеклассников в процессе изучения обществознания
- Использование сказки в коррекции отклонения в поведении младших школьников
- Эстетическое воспитание младших школьников
Поиск рефератов
Последние рефераты раздела
- Тенденции развития системы высшего образования в Украине и за рубежом: основные направления
- Влияние здоровьесберегающего подхода в организации воспитательной работы на формирование валеологической грамотности младших школьников
- Характеристика компетенций бакалавров – психологов образования
- Коррекционная программа по снижению тревожности у детей младшего школьного возраста методом глинотерапии
- Формирование лексики у дошкольников с общим недоразвитием речи
- Роль наглядности в преподавании изобразительного искусства
- Активные методы теоретического обучения