Углеводы
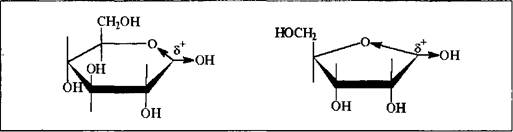
В соответствии с общими закономерностями реакций нуклеофильного замещения, в случае реализации мономолекулярного механизма ожидается образование продукта реакции в виде смеси диастереомеров, так как карбкатионный фрагмент переходного состояния молекулы
плоский; если же процесс пойдет по синхронному бимолекулярному пути - пространственная структура продукта будет зависеть от конфигурации исходного моносахарида: из б-формы образуется в-гликозид, из в-формы - б-гликозид, так как атака нуклеофила осуществляется в тыл связи С-0 и завершается обращением конфигурации реакционного центра. Поскольку моносахарид всегда будет существовать в растворе в виде таутомерной смеси а - и в-форм, то независимо от механизма реакции мы, как правило, получим гликозид в виде изомерной смеси. Только лишь их соотношение будет варьироваться в зависимости от условий реакций и природы реагента. Но это все верно тогда, когда реакция проводится в колбе, т.е. in vitro.
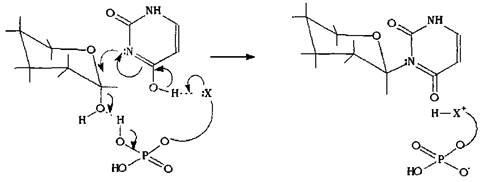
Внутри живой клетки такой процесс катализируется ферментами, которые, как правило, работают стереоспецифично: это значит, что они будут избирательно вовлекать в реакцию какой-либо один из диастереомеров, а также осуществлять реакцию по какому-либо одному механизму, что в итоге приведет к продукту одной изомерной формы. Схематично такую реакцию можно представить, моделируя реагент и фермент в виде единой молекулы, на одном конце которой находится остаток фосфорной кислоты, катализирующий отщепление гидроксильной группы, на другом конце - нуклеофильный остаток, атакующий электрофильный атом углерода. Синхронное воздействие каталитического и нуклеофильного фрагментов на электрофильный центр а-глюкопиранозы приводит к соответствующему в-гликозиду.
Вполне возможно, что б-изомер большинства моносахаров более реакционно способен хотя бы по причине стерической доступности его реакционного центра. Тогда следует ожидать, что большинство природных гликозидов будут относится к ряду в-изомеров, так как их образование ферментативно катализируемо. Трудно проанализировать
все природные гликозиды на предмет принадлежности их к а - или в-ряду в виду их многочисленности. Но все-таки создается впечатление, что в-диастереомеры все же более многочисленны. Так как б-изомерные формы часто также образуются исключительно стереоспецифично, следует предположить, что комплексирование каталитического сайта фермента и молекулы моносахарида чувствительно к исходной пространственной форме пиранозы или фуранозы. Это возможно только при условии, что образование одних и тех же гликозидов различной конфигурации должно катализироваться различными ферментативными системами.
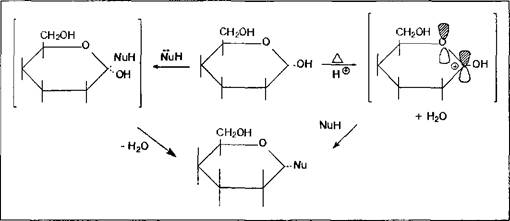
Для всех гликозидов характерно отсутствие таутомерных превращений в растворе, т.е. переход их в ациклическую форму невозможен, так как для этого процесса требуется подвижный атом водорода полуацетального гидроксила для преобразования последней в карбонильную группу. Второе общее свойство гликозидов - это достаточно легкая способность их к гидролизу в кислой среде, фактически являющемуся процессом, обратным реакции их образования.
Классификация гликозидов. Классифицируются гликозиды достаточно однозначно - согласно типу нуклеофила, заместившего полуацетальный гидроксил. Такой нуклеофил также называют агликоном. Если агликонами являются спирты или фенолы, то образующиеся гликозиды называют 0-гликозидами; серусодержащие нуклеофилы ведут к образованию S-гликозидов; если нуклеофилом являлось соединение, генерирующее в ходе реакции карбанион, то полученный гликозид имеет структуру С-гликозида; и наконец, очень распространены в природе различные варианты образования Н-гликозидов в ходе замещения полуацетального гидроксила моносахаров азотистыми основаниями разного типа.
О-гликозиды в природных источниках представлены как довольно простыми молекулами по структуре агликона, так и очень большими и разнообразными по строению и биологическим функциям соединениями, включая целые биополимерные системы.
Т. к. О-гликозиды образуются при взаимодействии спиртов любой природы с циклической формой моносахарида, а последний сам по себе содержит вполне достаточное количество различных по состоянию гидроксильных функций, то вполне естественно, что Природа использовала возможность образования новых структур, новых связей и новых возможностей путем взаимодействий молекул моносахаридов между собой по схеме О-гликозидирования, без привлечения других соединений. Такой путь всегда энергетически и материально наиболее экономичен.
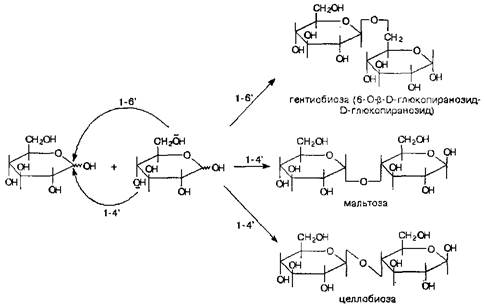
Результатом этих реакций являются дисахариды, олигосахариды и полисахариды, которые могут быть построены как из одинаковых моносахаридных звеньев, так и из различных моносахаридных звеньев.
Дисахариды. Самый важный момент, который следует определить в структуре дисахарида после того как установлена природа его моносахаридных звеньев - это характер гликозидной связи: какая гидроксильная группа участвует со стороны моносахарида - агликона и какова конфигурация гликозидной связи. Чаще всего реализуется связь 1-4', реже встречается гликозидная связь 1-6', еще реже - связь 1-3'.
Образованные таким способом дисахариды называются восстанавливающими, так как фрагмент моносахарида - агликона содержит полуацетальный гидроксил свободным, что оставляет за ним способность к таутомерному превращению в ациклическую форму, которая и вступит в окислительно-восстановительную реакцию своей альдегидной группой.
В том случае, если дисахарид образован связью 1-1', т.е. моносахарид - агликон предоставляет для формирования гликозидной связи свой полуацетальный гидроксил - его относят к группе невосстанавливающих, так как таутомерного превращения, ведущего к ациклическому фрагменту с альдегидной группой, молекула не претерпевает и, следовательно, легких окислительно-восстановительных реакций от нее ожидать не приходится.
Типичными представителями гетеродетных дисахаридов являются лактоза и сахароза. Само название этих дисахаридов говорит о степени важности их для живых организмов и степени распространенности в живой системе. Лактоза представляет собой дисахарид, образованный из галактозы и глюкозы-агликона со связью в-1-4", тогда как сахароза образована из глюкозы и фруктозы со связью 1б-1'в, т.е. из этих данных следует, что лактоза является восстанавливающим дисахаридом, а сахароза – не восстанавливающим.
Полисахариды также должны быть классифицированы как О-гликозиды, поскольку образуются точно по той же схеме и имеют ту же природу связывания моносахаридных звеньев между собой Само собой разумеется, что от О-гликозидов дисахаридного типа они отличаются количеством этих звеньев - многие природные полисахариды имеют весьма высокую степень полимерности. Поскольку полисахариды, как и все биополимеры - это все-таки больше объекты биохимии и химии высокомолекулярных соединений, мы приведем здесь лишь краткое описание основных типов макромолекул этого класса веществ.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики