Действие и синтез гормонов
а) Г + Р + Gи·ГДФ ® Г·Р· Gи + ГДФ б) Г·Р·Gи + ГТФ ® Г + Р + Gи·ГТФ в) Gи·ГТФ + А ® ЇцАМФ + Gи·ГДФ
После фосфорилирования белков-ферментов в ходе выше описанных реакций (см. рис 1) изменяется их конформация. Следовательно изменяется и конформация их активного центра, что ведет к их активированию или ингибированию. Получается, что благодаря вторичному посреднику цАМФ в клетке активируются или
ингибируется действие специфичных для нее ферментов, что вызывает определенный биологический эффект свойственный для этой клетки. В связи с этим, несмотря на большое количество ферментов, которые действуют через вторичный посредник цАМФ, в клетке возникает определенный, специфический ответ.
Кальмодулин
Эффект кальмодулина очень похож на эффект цАМФ. В клетке по мере дифференцировки образуется набор Са2+-кальмодулин-чувствительных белков. Кальмодулин активирует ферменты связываясь с ними и изменяя их конформацию. В то же время кальмодулин сам является аллостерическим белком, который проявляет свои биологические свойства только в связанном с ионами Са виде. Комплекс кальмодулина с Са может изменять активность белков одним из двух способов:
1) путем прямого воздействия на фермент-мишень
2) через активируемую этим комплексом протеинкиназу
Взаимодействие вторых посредников
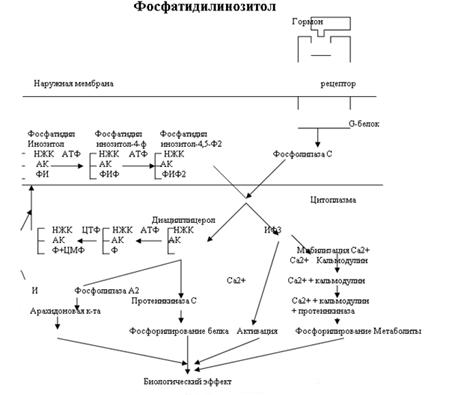
Как было показано на рис.2 Фосфатидилинозитол также запускает кальмодулин. В то же время кальмодулин как и цАМФ активируют белки-ферменты благодаря чему вызывается биологический эффект. Даже среди 3 известных на сегодняшний день типов вторичных посредников мы видим тесное взаимодействие.
Рассмотрим взаимодействие кальмодулина и цАМФ. Большое разнообразие набора кальмодулиновых белков в клетке ведет к различному действию на цАМФ в разных клетках организма. В одном случае кальмодулин активирует фосфодиэстеразу (см. рис.1) что приводит к ингибированию эффекта цАМФ. В другом случае он активирует аденилатциклазу, в третьем ингибирует. В связи с таким разнообразием эффектов Расмуссен предположил 5 различных ситуаций, в которых конечная интегральная реакция зависит от одновременного присутствия цАМФ и Са2+, действующих вместе или последовательно:
· равноправное партнерство, когда оба посредника необходимы для полного проявления биологического эффекта
· тот или другой посредник играет главную роль, а другой лишь облегчает его роль
· посредники действуют последовательно, т.е. высвобождение Са повышает продукцию цАМФ (активирует аденилатциклазу) и дальше они действуют вместе
· двойной контроль является излишним
· посредники являются антагонистами, т.е. выступают в роли «сигнала выключения друг друга»
Гормон роста (соматотропин)
Как и каждый гормон аденогипофиза соматотропин синтезируется под действием рилизинг фактора (ГР-РФ). Стимуляторами выработки ГР-РФ являются тиреоидный гормоны и глюкокортикоиды. Главным ингибитором реактивности питуицитов по отношению к ГР-РФ является соматостатин. Он стимулируется соматомединами. То есть можно сказать что соматостатин действует по методу обратной связи.
ГР-РГ стимулирует как секрецию так и синтез СТГ. Этот эффект опосредуется связыванием ГР-РФ с рецепторами плазматической мембраны соматотрофов и активацией трех систем вторичных посредников. С их помощью происходит усиленный синтез мРНК СТГ. Этот эффект действия водорастворимого гормона на ядерный аппарат клетки объясняют фосфорилированием и дефосфорилированием определенной группы протеинкиназ, которые действуют на генетический аппарат клетки. На данный момент ни одна из этих протеинкиназ не идентифицирована.
СТГ не действует прямо на клетку. Он переносится кровью в печень где превращается в соматомедины. То что СТГ не действует прямо на гормон было доказано на опыте, когда изолированную хрящевую ткань перфузировали раствором, который содержал большое количество СТГ и не наблюдали никакого ее роста. При помещении ее в нормальную сыворотку крови ее клетки росли. Потом были найдены соматомедины, которые непосредственно вызывают эффект роста.
Клеточный механизм действия СТГ на хрящевую ткань связан с эффектами соматомедина. Хотя стимуляция роста и других тканей, таких как печень и мышцы, может опосредоваться аналогичным механизмом, in vitro показано и прямое действие СТГ на мышцу. Так, подобно инсулину СТГ вызывает независимое от синтеза белка усиление транспорта глюкозы. Спустя некоторое время после добавления СТГ мышца приобретает резистентность к действию инсулина, и в этот период усиливается транспорт аминокислот, зависящий от синтеза нового белка. Действие СТГ на печень до сих пор недостаточно изучено. Гормон стимулирует синтез белка, РНК. Это в свою очередь ведет к увеличению количества полисом. Также в конце концов он стимулирует синтез ДНК. На данный момент неизвестно, как СТГ, имеющий рецепторы на мембране может вызывать эффекты похожие на действие стероидных гормонов. Можно предположить, что он вызывает такие эффекты вызывая синтез ферментов, отвечающих за синтез нуклеиновых кислот. Это также доказывает то, что жирорастворимые гормоны включают только определенные гены, а СТГ ведет к генерализованой реакции увеличения синтеза белка на включенных генах, что ведет к росту клетки, а не к ее дифференциации.
До сих пор не вполне ясно, какие эффекты вызывает непосредственно СТГ, а какие соматомедины. Известно точно, что соматомедины действуют непосредственно на хрящевую ткань, вызывая в хондробластах:
· стимуляцию включения SO4 в протеогликаны
· стимуляцию включения тимидина в ДНК
· стимуляцию синтеза РНК
· стимуляцию синтеза белка
Тиреоидные гормоны (Т3 – Т4)
Пептидергические нейроны в преоптической области гипоталамуса синтезируют и выделяют в воротную систему гипофиза тиреотропин-рилизинг гормон (ТРГ). ТРГ стимулирует выработку в клетках гипофиза тиреотропного гормона (ТТГ). ТТГ представляет собой гликопротеид с молекулярной массой 29000. ТТГ может влиять на развитие тиреоидных фолликулярных клеток, предшествующих образованию самих фолликулов в щитовидной железе. Так, было показано, что тиреоидные клетки in vitro, диспергированные с помощью протеолитических ферментов, при добавлении в среду ТТГ вновь собираются в молекулы. Это показывает, что ТТГ способствует синтезу каких-то компонентов клеточной мембраны, необходимых для распознавания друг друга. ТТГ-рецепторный комплекс, образующийся в результате связывания ТТГ, вызывает ответные биологические реакции с помощью не менее четырех внутриклеточных посредников: цАМФ, инозитолтрифосфата, диацилглицерола и комплекса Са2+-кальмодулин. Интересно, что активация аденилатциклазы в мембранах тиреоидных клеток под действием ТТГ в основном обуславливается ингибированием Gи-белка. Под влиянием ТТГ в клетках тиреоидных фолликулов возникают глубокие морфологические изменения. Тиреоидная клетка в высокой степени полярна: на ее апикальной поверхности имеются многочисленные микроворсинки (увеличение площади), к которым прилегают запасы коллоида в фолликуле. Базальная поверхность клетки контактирует с кровью. Коллоид представляет собой белковый раствор богатый на остаток аминокислоты тирозина. В ходе биохимических реакций фенольный гидроксид тирозина соединяется с фенолом другого тирозина и присоединяя к себе йод образуется тетрайодтиронин, связанный пептидными связями с остатками других аминокислот. Такие белки называются тиреоглобулинами.
Другие рефераты на тему «Медицина»:
Поиск рефератов
Последние рефераты раздела
- Особенности лечения и тракционно-экстензионной терапии на аппарате Kinetrac KNX-7000
- Остеохондроз, методики лечения
- Тракционно-экстензионная терапия у больных остеохондрозом пояснично-крестцового отдела позвоночника
- Болезни, возникающие от курения. Профилактика курения
- Болезни органов дыхания
- Болезни желчевыводящих путей и печени
- Анатомия и физиология артерий нижних конечностей. Этиология и патогенез