Болевой анализатор
Сенситизация. Общепризнано, что большинство типов сенсорных рецепторов становятся менее чувствительными, если подвергаются длительной стимуляции. В отличие от них у ноцицепторов чувствительность прогрессивно возрастает (до определенных пределов), если действие сохраняется продолжительное время. Такое снижение пороговой чувствительности называется сенситизацией. Это явление обнаружено так
же в процессе регенерации поврежденного нервного ствола, когда разрастающаяся соединительная ткань вызывает сжатие регенерирующих тонких нервных волокон. При этом в нерве регистрируется необычайно высокая фоновая ритмика и низкий порог для активации волокон в области повреждения. Согласно одной из гипотез о происхождении этого феномена, повреждение тканей вызывает высвобождение рецепторной субстанции, сопровождающееся инактивацией разрушающего ее фермента и стойкой деполяризацией мембраны рецептора, что обусловливает устойчивые разряды в афферентном волокне. Сенситизация проявляется в способности ноцицептора отвечать на адекватные стимулы допороговой величины (например, активироваться при температуре 40 °С, а не 45 °С, как отмечалось ранее), а также в способности возбуждаться ноцицептивными стимулами прежде неэффективных модальностей. Явление сенситизации многие исследователи рассматривают как одну из причин кожной гиперальгезии.
Центральные системы боли
Цитоархитектоника задних рогов спинного мозга. В дорсальных рогах спинного мозга находятся центральные нейроны, воспринимающие и перерабатывающие болевые сигналы перед отправкой их в головной мозг. Задний рог принято делить на 5 – 6 пластин. Первая пластина – краевая зона, содержит маргинальные нейроны, которые являются ноцицептивными интернейронами спиноталамического пути. Вторая – третья пластины образуют тесное сплетение мелких нейронов – желатинозную субстанцию, содержащую в себе малые тормозящие интернейроны, создающие сложные замкнутые цепочки, и крупные нейроны, аксоны которых идут в спиноталамический путь. Важной структурой желатинозной субстанции являются гломерулы – сложные синапсы между терминалями первичных афферентных волокон, дендритами центральных нейронов и окончаниями нисходящих путей. Эти структуры, как полагают, обеспечивают взаимодействие между афферентными и центральными потоками импульсов в процессе переработки информации. Четвертая и пятая пластины, или собственное ядро, - зона скопления самых крупных нейронов заднего рога, дендриты которых проникают в желатинозную субстанцию, а аксоны многих из них формируют спиноталамический путь. Нейроны собственного ядра легко активируются болевыми и неболевыми стимулами. Более глубокие пластины серого вещества (седьмая – восьмая), не относящиеся к заднему рогу, дают начало спиноретикулярным волокнам.
Центральные проекции первичных афферентных волокон. Перед входом в спинной мозг толстые и тонкие волокна дорсальных корешков перераспределяются. При этом тонкие миелиновые и немиелиновые волокна, несущие информацию о боли, группируются в отдельные пучки, которые входят в спинной мозг и образуют зону Лиссауэра. Пройдя в ней вдоль спинного мозга 1 – 2 сегмента, терминали волокон проникают в первую пластину серого вещества задних рогов. Толстые миелиновые волокна, войдя в спинной мозг, образуют петлю, входят в серое вещество с внутренней стороны задних рогов через пятую пластину и лишь затем достигают желатинозной субстанции (вторая – третья пластины).
Тонкие первичные афферентные волокна оканчиваются в первой половине – маргинальной зоне, а большинство толстых афферентных волокон – во второй пластине желатинозной субстанции. Перед окончанием толстые афферентные волокна проходят вдоль внутренней поверхности задних рогов, входят в серое вещество через четвертую – пятую пластины и затем поднимаются во вторую пластину. Здесь они оканчиваются факелоподобными пучками, но не проникают в маргинальный слой. Факелоподобные пучки окончаний толстых афферентных волокон делят желатинозную субстанцию на дольки, в которых нейроны организованы в вертикальные столбы. Крупные нейроны четвертой пластины посылают свои дендриты в эти дольки, где и устанавливаются синаптические контакты с толстыми первичными афферентными волокнами.
Считается доказанным, что нейроны второй – третьей пластин (область желатинозной субстанции) во взаимодействии с маргинальными нейронами образуют специализированную систему модулирующего контроля болевого афферентного входа, а пятая пластина является уровнем полимодального ноцицептивного входа и областью взаимодействия с толстыми неболевыми афферентами (см. рис. 2).
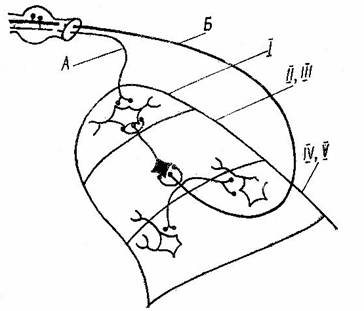
Рис. 2 Схема некоторых связей толстых и тонких первичных афферентов нейронами заднего рога.
А – тонкие, Б – толстые афференты, Й – маргинальный слой, ЙЙ и ЙЙЙ – желатинозная субстанция, ЙV и V – собственное ядро.
Тонкие афферентные волокна от кожи, мышц, сухожилий и внутренних органов (группы А-۵-, С, ЙЙ, ЙЙЙ, ЙV) через интернейроны заднего рога замыкается на мотонейронах сгибательных мышц и образуют обширную группу сегментарных сгибательных рефлексов, которые играют важную роль в ноцицептивных механизмах, создавая защитные рефлекторные реакции.
Функциональная организация нейронных цепей в заднем роге. В настоящее время предложены две основные схемы взаимодействия афферентных сигналов в заднем роге. Вопрос о входе к маргинальным нейронам пока не решен. Зона Лиссауэра содержит не только первичные афферентные волокна, но и аксоны нейронов желатинозной субстанции. После перерезки этой зоны аксосоматические и аксодендритные синапсы на маргинальных нейронах дегенерируют, что указывает на то, что С-волокна и аксоны нейронов желатинозной субстанции должны оканчиваться на них. Как уже упоминалось, С-волокна оканчиваются в маргинальной зоне и желатинозной субстанции. В свою очередь, нейроны желатинозной субстанции образуют тормозящие синапсы на соме маргинальных нейронов.
Толстые афферентные волокна заканчиваются возбуждающими синапсами на нейронах желатинозной субстанции и больших нейронах собственного ядра. Полагают, что ритмическая активность нейронов желатинозной субстанции импульсами от толстых афферентных волокон может выражаться в интенсивном торможении маргинальных нейронов, на которых образуют свои синапсы тонкие волокна. Такая нейронная цепь может создать механизм, посредством которого толстые неболевые афферентные волокна моделируют порог боли. На рисунке показана принципиальная схема возможной организации такой системы контроля.
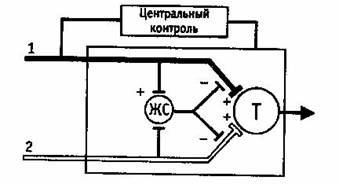
Рис.3 Схема контроля интенсивности сенсорного потока в заднем роге серого вещества спинного мозга.
А – тонкие, Б – толстые афференты, ЖС – желатинозная субстанция, Т – нейроны, передающие сенсорную информацию в мозге, (+) – возбуждение, (-) – торможение.
Толстые неболевые афференты и тонкие болевые афференты подходят к нейронам в желатинозной субстанции и центральным нейронам, передающим сенсорную информацию в мозг (Т-клетки). Тормозящий эффект от нейронов желатинозной субстанции, возникающий в аксоаксональных синапсах на толстых и тонких афферентах, увеличивается, если возрастает поток импульсации в толстых неболевых афферентах, и уменьшается, если повышается поток импульсации по тонким болевым афферентам. Существуют также механизмы центрального контроля, которые управляются потоком импульсации по толстым афферентам. Эти механизмы проецируются в воротную систему контроля, Т-клетки посылают сигналы в систему действия.
Другие рефераты на тему «Медицина»:
Поиск рефератов
Последние рефераты раздела
- Особенности лечения и тракционно-экстензионной терапии на аппарате Kinetrac KNX-7000
- Остеохондроз, методики лечения
- Тракционно-экстензионная терапия у больных остеохондрозом пояснично-крестцового отдела позвоночника
- Болезни, возникающие от курения. Профилактика курения
- Болезни органов дыхания
- Болезни желчевыводящих путей и печени
- Анатомия и физиология артерий нижних конечностей. Этиология и патогенез