Анатомия центральной нервной системы
а - афферентные волокна; б - типы клеток, выявляемые на препаратах, импрегнированных по методу Голдбжи; в - цитоархитектоника, выявляемая при окрашивании по Нисслю. 1 - горизонтальные клетки, 2 - полоска Кеса, 3 - пирамидные клетки, 4 – звездчатые клетки, 5 - наружная полоска Белларже, 6 - внутренняя полоска Белларже, 7 - видоизмененная пирамидная клетка.
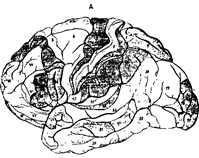
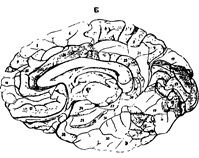
Рис. 37. Цитоархитектоника (А) и ми
елоархитектоника (Б) коры полушарий большого мозга.
При сохранении общего плана строения было установлено, что разные участки коры (в пределах одной области) отличаются по толщине слоев. В некоторых слоях можно выделить несколько подслоев. Кроме того, имеются различия клеточного состава (разнообразие нейронов, плотность и их расположение). С учетом всех этих различий Бродман выделил 52 участка, которые назвал цитоархитектоническими полями и обозначил арабскими цифрами от 1 до 52 (рис.38 А,Б ).
А вид сбоку. Б срединно-сагиттальный; срез.
Рис. 38. Схема расположения полей по Бордману
Каждое цитоархитектоническое поле отличается не только клеточным строением, но и расположением нервных волокон, которые могут идти как в вертикальном, так и в горизонтальном направлениях. Скопление нервных волокон в пределах цитоархитектонического поля называют миелоархитектоникой.
В настоящее время все большее признание находит “колончатый принцип” организации проекционных зон коры.
Согласно этому принципу каждая проекционная зона состоит из большого количества вертикально ориентированных колонок, диаметром приблизительно в 1 мм. Каждая колонка объединяет около 100 нейронов, среди которых имеются сенсорные, вставочные и эфферентные нейроны, связанные между собой синаптическими связями. Одиночная “корковая колонка” участвует в переработке информации от ограниченного количества рецепторов, т.е. выполняет специфическую функцию.
Система волокон полушарий
Оба полушария имеют три типа волокон. По проекционным волокнам возбуждение поступает в кору от рецепторов по специфическим проводящим путям. Ассоциативные волокна связывают между собой различные области одного и того же полушария. Например, затылочную область с височной, затылочную – с лобной, лобную – с теменной областью. Комиссуральные волокна связывают симметричные участки обоих полушарий. Среди комиссуральных волокон выделяют: переднюю, заднюю мозговые спайки и мозолистое тело ( рис. 39 А.Б).
|
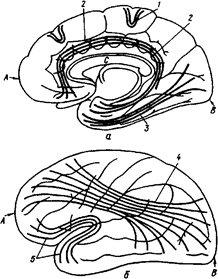
Рис. 39А. а - медиальная поверхность полушария;
б - верхнеальтеральная поверхность полушария;
А - лобный полюс;
В - затылочный полюс;
С - мозолистое тело;
1 - дугообразные волокна большого мозга соединяют между собой соседние извилины;
2 - пояс - пучок обонятельного мозга лежит под сводчатой извилиной, простирается из области обонятельного треугольника до крючка;
3 - нижний продольный пучок связывает затылочную и височную область;
4 - верхний продольный пучок связывает лобную, затылочную, височную доли и нижнетеменную дольку;
5 - крючковидный пучек расположен у переднего края островка и соединяет лобный полюс с височным.
Рис. 39Б. Кора головного мозга на поперечном разрезе. Оба полушария соединены пучками белого вещества, образующими мозолистое тело (комиссуральные волокна).
Рис. 39. Схема ассоциативных волокон
Ретикулярная формация
Ретикулярная формация (сетчатое вещество мозга) была описана анатомами в конце прошлого века.
Ретикулярная формация начинается в спинном мозге, где она представлена желатинозной субстанцией основания заднего мозга. Основная ее часть находится в центральном стволе мозга и в промежуточном мозге. Она состоит из нейронов различной формы и размеров, которые имеют обширные ветвящиеся отростки, идущие в разных направлениях. Среди отростков выделяют короткие и длинные нервные волокна. Короткие отростки обеспечивают локальные связи, длинные - формируют восходящие и нисходящие пути ретикулярной формации.
Скопления нейронов образуют ядра, которые находятся на разных уровнях мозга (спинном, продолговатом, среднем, промежуточном). Большинство ядре ретикулярной формации не имеют четких морфологических границ и нейроны этих ядер объединяются только по функциональному признаку (дыхательный, сердечно-сосудистый центр и др.). Однако на уровне продолговатого мозга выделяют ядра с четко обозначенными границами - ретикулярное гигантоклеточное, ретикулярное мелкоклеточное и латеральное ядра. Ядра ретикулярной формации моста по существу являются продолжением ядер ретикулярной формации продолговатого мозга. Наиболее крупные из них - каудальное, медиальное и оральное ядра. Последнее переходит в клеточную группу ядер ретикулярной формации среднего мозга и ретикулярное ядро покрышки мозга. Клетки ретикулярной формации являются началом как восходящих, так и нисходящих путей, дающих многочисленные коллатерали (окончания), которые образуют синапсы на нейронах разных ядер центральной нервной системы.
Волокна ретикулярных клеток, направляющихся в спинной мозг, образуют ретикулоспинальный тракт. Волокна восходящих трактов, начинающихся в спинном мозге, связывают ретикулярную формацию с мозжечком, средним мозгом, промежуточным мозгом и корой полушарий большого мозга.
Выделяют специфические и неспецифические ретикулярной формации. Например, некоторая часть восходящих путей ретикулярной формации получают коллатерали от специфических путей (зрительных, слуховых и т.д.), по которым афферентные импульсы передаются в проекционные зоны коры.
Неспецифические восходящие и нисходящие пути ретикулярной формации влияют на возбудимость различных отделов мозга, в первую очередь коры полушарий большого мозга и спинной мозг. Эти влияния по функциональному значению могут быть как активирующими, так и тормозными, поэтому выделяют: 1) восходящее активирующее влияние, 2) восходящее тормозное влияние, 3) нисходящее активирующее влияние, 4) нисходящее тормозное влияние. На основании этих факторов ретикулярную формацию рассматривают как регулирующую неспецифическую систему мозга.
Наиболее изучено активирующее влияние ретикулярной формации на кору полушарий большого мозга. Большинство восходящих волокон ретикулярной формации диффузно оканчивается в коре полушарий и поддерживает ее тонус, обеспечивает внимание. Примером тормозных нисходящих влияний ретикулярной формации является снижение тонуса скелетных мышц человека во время определенных стадий сна.
Нейроны ретикулярной формации чрезвычайно чувствительны к гуморальным веществам. Это опосредованный механизм влияния различных гуморальных факторов и эндокринной системы на высшие отделы мозга. Следовательно, тонические воздействия ретикулярной формации зависят от состояния всего организма (рис.40).
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики