Спецификация нервной ткани
Региональная спецификация нервной ткани
У взрослых животных клетки различных областей нервной системы имеют существенные отличия в своем фенотипе в зависимости от той функции, которую они выполняют. Клетки мозжечка отличаются от клеток коры полушарий, и оба типа клеток — от сетчатки. Каким образом происходит формирование таких различных фенотипов клеток в процессе развития? Как и у
других клеток, фенотип нейрона определяется тем, какие гены он экспрессирует, что, в свою очередь, контролируется факторами транскрипции, белками, которые связываются с регуляторными зонами одного или нескольких генов и влияют на процессы их транскрипции. Развитие характеризуется последовательной и иерархической экспрессией факторов транскрипции, каждый из которых влияет на экспрессию последующего и ограничивает конечный фенотип клетки.
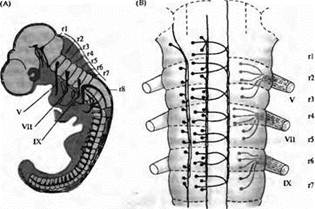
Рис. 1. Развитие заднего мозга позвоночных β виде сильно сегментированной структуры. (А) Диаграмма трехдневного эмбриона цыпленка, иллюстрирующая сегментарную организацию ромбомеров (r1-r8) в заднем мозге. (В) Паттерн организации клеток в ромбомерах r1-r7 трехдневного мозга цыпленка. Ретикулярные нейроны (слева) и брахиомоторные нейроны (справа) образуют сегментарный повторяющийся паттерн. Мотонейроны и их аксоны образуют черепно-мозговые нервы V, VII и IX пары.
Исследованию региональной спецификации нервной ткани в мозгу позвоночных значительно помогло то, что были открыты гомологи генов плодовой мухи дрозофилы, которые определяют расположение и зависящую от расположения дифференцировку клеток эмбриона. Эти гены оказались сходными с генами позвоночных и часто выполняют сходные функции. Большинство из этих генов кодирует факторы транскрипции.
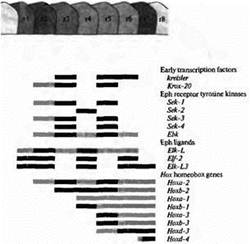
Рис. 2. Сегментарная экспрессия генов в ромбомерах с r1 до r8 в заднем мозге позвоночных. Серые полоски указывают ромбомеры, в которых происходит экспрессия генов; черные полоски указывают высокий уровень экспрессии. Ранние факторы транскрипции, рецептор тирозинкиназы семейства Eph и его лиганды образуют сегментарный паттерн ромбомеров Семейство гомеобоксных генов Нох определяет судьбу клеток в пределах каждого ромбомера в зависимости от сегмента. Данные получены на эмбрионах цыплят и мышей.
Гомеотические гены и сегментация
Удивительными примером подобного сходства в функционировании генов у настолько разных животных являются результаты исследований заднего мозга позвоночных. В отличие от всего остального мозга позвоночных, задний мозг эмбриона (rhombencephalon) имеет четкое сегментарное строение. Каждый сегмент его демонстрирует общий паттерн нервной дифференцировки, однако от сегмента к сегменту дифференцировка имеет свои особенности (рис. 1). Было идентифицировано несколько генов, паттерны экспрессии которых на ранних стадиях развития коррелируют с границами сегментов заднего мозга (рис. 2). Эти гены делятся на две категории:
(1) Гены первой категории играют роль в создании общей структуры, состоящей из повторяющихся сегментных единиц. Некоторые гены этой группы кодируют факторы транскрипции (kreisler, Krox-20), другие кодируют рецептор тирозинкиназы (Sek-l по Sek-4) или его лиганды (Elf-2). (Рецептор тирозинкиназы представляет собой трансмембранный белок, у которого внутриклеточный домен, представляющий собой фермент тирозинкиназу, активируется при связывании лиганда с внеклеточным доменом.)
(2) Вторая категория содержит гены, которые определяют судьбу каждого сегмента. Эти гены образуют высококонсервативное семейство Нох генов.
Свойства Нох генов были впервые описаны у дрозофилы, у которой они регулируют гомеостаз. Гомеотические гены представляют собой управляющие гены, которые координируют экспрессию многих других генов во время развития. Например, мутация гомеотических генов семейства Нох у дрозофилы приводит к тому, что одна часть тела заменяется другой; таким образом на месте антенны может развиться нога. Гомеотические гены содержат консервативную последовательность ДНК, гомеобокс. Гомеобокс кодирует последовательность из 60 аминокислот, которые распознают и связываются с определенной последовательностью ДНК серии подчиненных генов. Каждый гомеотический ген, таким образом, координирует экспрессию большого числа генов, которые вместе определяют строение одного сегмента эмбриона.
Сегментарный паттерн экспрессии генов семейства Нох наблюдается в заднем мозгу цыплят и грызунов, что приводит нас к предположению, что Нох гены могут играть роль гомеотических управляющих генов, регулирующих процессы развития позвоночных и создания определенных структур относительно рострокаудальной оси в определенных областях заднего мозга эмбриона. Данные, полученные при помоши трансплантации, выключения определенных генов и эктопической экспрессии, согласуются с этими идеями. Дополнительным доказательством является изучение мутаций Нох генов и других гомеобоксных генов у человека, которые приводят к изменениям в определенных областях ЦНС.
Следующий очевидный вопрос: а что определяет паттерн экспрессии Нох генов? Ответом, по крайне мере частично, является градиент ретиноевой кислоты. Ретиноевая кислота вырабатывается в Гензеновском узелке, который называется Шпемановским организатором (Spemann organizer) у эмбрионов птиц и позвоночных (см. рис. 3). Но ретиноевая кислота не просто активирует транскрипцию всех Нох генов: была описана систематическая разница чувствительности различных генов Нох семейства к ретиноевой кислоте. Таким образом, диффузия ретиноевой кислоты из Гензеновского узелка способствует формированию градиента, который играет большую роль в упорядоченной экспрессии различных Hax генов в рострокаудальном направлении в заднем мозге.
Хорда и базальная пластинка
Строение и функпчонирование нервной системы позвоночных варьируют в дорзовентральном и рострокаудальном направлении. Например, полоска специализированных глиальных клеток, названных базальной пластинкой (floor plate), расположена по средней линии вдоль вентральной поверхности спинного мозга. Соседние, более латерально расположенные базальиые области нервной трубки образуют мотонейроны, более дорзальные зоны дают начало интернейронам, и самые дорзальные области образуют нервный гребень.
Характерные свойства вентральной хорды, такие как дифференцировка базальной пластинки и образование мотонейронов, регулируются сигналом из спинного мозга (notochord). Таким образом, если пересадить в эмбрион еше одну хорду, рядом с нервной трубкой, то это приведет к формированию второй базальной пластинки и второй группы мотонейронов (рис. 3), а если у эмбриона удалить хорду, то ни мотонейроны, ни базальная пластинка не формируются.
Сигналы из хорды, которые управляют формированием клеток базальной пластинки и мотонейронов, являются продуктами транскрипции гена Sonic hedgehog24). Белок Sonic hedgehog синтезируется клетками хорды (а затем также клетками базальной пластинки), концентрируется на их поверхностях и диффундирует к соседним клеткам. Высокий уровень Sonic hedgehog на поверхности хорды приводит к формированию клеток базальной пластинки из клеток нервной трубки. Более низкий уровень ведет к экспрессии гомеотического гена (Mkx-2.2), что вызывает развитие клеток в висцеральные мотонейроны. Еще более низкий уровень Sonic hedgehog вызывает транскрипцию Рах-6, которые угнетают экспрессию Nkx-2.2, что позволяет клеткам развиваться по их основному пути и становиться соматическими мотонейронами.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики