Сенсорные системы живых организмов
Эхолокация
Эхолокация состоит в определении пространственного положения объекта благодаря отражению этим объектом звуковых сигналов, излучаемых самим животным-наблюдателем. При этом животные, обладающие эхолокационным механизмом, опознают этим способом размеры, форму и материал объектов.
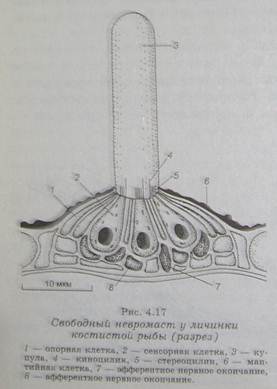
Рис.1
Эхолокация
развилась у тех животных, которые по крайней мере часть своей активной жизни проводят в таких условиях, где функции зрительного анализатора ограничены или полностью исключаются (летучие мыши, крыланы, стрижи-саланганы, гуахаро).
Развитию специализированных сонаров предшествовало первоначально чувство препятствия (обнаруживается у слепых людей, когда они подсознательно воспринимают звуки, отражающиеся от предметов), следующим этапом развития была неспециализированная эхолокация, заключающаяся в рефлекторном озвучивании пространства.
Самыми примитивными являются сонары пещерных птиц, представителей рода летучих собак из семейства крылановых и ушастых тюленей. Все они используют в качестве локационных сигналов широкополосные щелчки, основная энергия которых у птиц сосредоточена в определенном диапазоне частот. Щелчки издаются механическим способом – прищелкивание клювом или языком. Звуковое частотное заполнение сигналов обуславливает низкую разрешающую способность их сонаров, которые, выполняют функцию обнаружения препятствия и оценку расстояния до него.
Наибольшего совершенства эхолокационная функция достигла у подотряда летучих мышей и зубатых китообразных. Качественное отличие от эхолокации птиц и крыланов заключается в использовании ультразвука. Малая длина волны ультразвуковых колебаний создает благоприятные условия для получения четких отражений даже от небольших предметов, которые волны слышимого диапазона огибают. Кроме того, ультразвук можно излучать узким, почти параллельным пучком, что позволяет концентрировать энергию в нужном направлении. В формировании локационных сигналов у летучих мышей и зубатых китов участвуют специализированные гортанные механизмы и система носовых мешков, а в качестве каналов для излучения ультразвука используются ротовая и носовая полости, а так же специализированный лобный выступ – мелон.
Таким образом, возникновение эхолокации стало возможным лишь после освоения животными трехмерного пространства (воздушной или водной среды) в таких экологических условиях, где оптическими средствами невозможно получить какую-либо информацию о наличии препятствий (пещеры, подводный мир).
Сенсорная система боковой линии
Основной структурной адаптацией боковой линии является эпидормальный орган – невромаст. Невромасты могут располагаться на поверхности тела у водных амфибий и рыб (свободные) или внутри системы каналов боковой линии у большинства рыб (канальные). Каналы заполнены жидкостью или слизистым содержанием и контактируют с внешней средой через поры, закрытые у некоторых видов мембраной.
Каждый невромаст содержит группу волосковых клеток, расположенных среди опорных клеток. На апикальной поверхности волосковой клетки находится набор сенсорных волосков (цилий) – 30-40 стереоцилий и единичная эксцентрично расположенная киноцилия. На противоположной, базальной поверхности волосковая клетка образует синаптические контакты с нервными окончаниями, которые могут быть аффективными и эффективными, что позволяет рассматривать эти клетки как типичные вторичночувствующие рецепторные образования. С поверхности волосковых клетов цилии вдаются в кУпулу из желатинообразного вещества, которая лежит над сенсорной областью каждого невромаста. Купулы свободных нефромастов, например, у шпорцевой лягушки, часто имеют удлиненную форму и выдаются в водную среду, купулы канальных невромастов с овальными дискообразными сенсорными полями, как у налима, часто представлены сводчатой формой.
Таким образом, у свободных невромастов вся вспомогательная структура рецепторного аппарата ограничена купулой, тогда как у канальных между рецептирующей волосковой клеткой могут находиться не только вещество купулы, но еще и структуры, образующие стенку канала, а так же жидкость, заполняющая этот канал с повышенным содержанием калия. Слизистые купулы содержат сеть микрофибрилл и приклепляются ими к цилии волосковой клетки.
Ответ волосковой клетки вызывается усилением сдвига, создаваемого движением основания купулы, где расположены цилии. Микрофибриллы,связанные с цилиями, передают этим сенсорным структурам информациюо степени смещения. В результате возникающее движение вызывает градуальный потенциал волосковой клетки.
Свободные невромасты рыб и амфибий стимулируются преимущественно струями воды, омывающими поверхность тела. Канальные невромасты в большей или меньшей степени изолированы от этих раздражений и восприимчивы к гидродинамическим полям и инфразвуковым вибрациям. Установлено, что волосковые клетки реагируют на смещение водных частиц, вызываемых близлежащими источниками колебаний. У некоторых видов они так же приобретают высокую чувствительность к температуре и солености воды.
Восприятие электрических сигналов
По способности воспринимать и использовать в своей жизнедеятельности электрические поля водные животные разделяются на 2 группы. У большинства рыб – неэлектрочувствительные виды – реакции на электрическое поле проявляются при градиенте потенциала более 1мВ/см. Эти реакции носят, как правило, непроизвольный характер и не связаны с наличием специфических электрорецепторов.

Рис.2
У ряда животных в процессе эволюции возникли высокочувствительные сенсорные системы, предназначенные для восприятия электрических сигналов в водной среде. Впервые существование таких органов было предсказано в 1951 г. Г.Лиссманом. Схематически процесс электровосприятия, по Лиссману, состоит в том, что в отсутствие раздражителя животное окружено электрическим полем определенной конфигурации. Оно создается непрерывно работающим электрическим органом, расположенным у основания хвоста. Это поле воспринимается электрорецепторами боковой линии. Объект, отличны по электропроводности от окружающей воды, искажает поле рыба и изменяет активность электрорецепторов.
Дальнейшие исследования подтвердили существование у ряда первичноводных позвоночных специализированных электрорецепторов, которые по строению и функциям отличаются от обычных механорецепторов боковой линии. Электрорецепторные сенсорные клетки, так же как и волосковые клетки невромастов, образуют синаптические контакты с подходящими к ним волокнами нерва боковой линии, т.е. тоже являются вторичночувствующими. Однако на базальной поверхности у них располагаются только аффетентные синапсы; эфферентной иннервации не обнаружено. Синаптические контакты имеют вид ленты, которая проникает вглубь рецепторной клетки на значительное расстояние. На одной клетке может быть несколько синапсов, а одно и то же нервное волокно способно контактировать сразу с сотнями соседних клетов.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики