Вопросы экологии применительно к лесному хозяйству
Повышение популяцией
потенциальной емкости
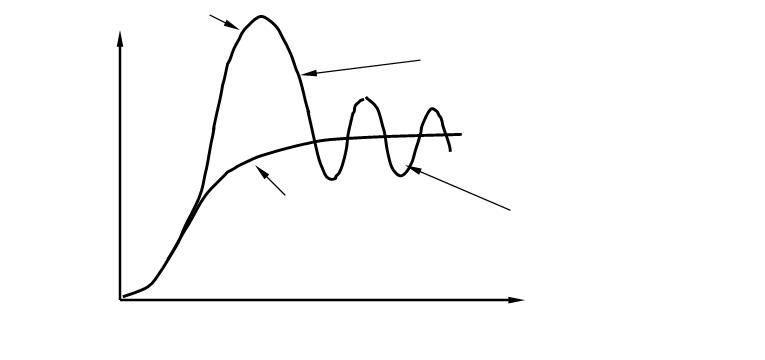
«Крах» популяции
или вымирание
Число
особей
Потенциальная Популяционные
емкость волны
Время
Рис. 4. Преобразование J –образной кривой роста численности популяции в S – обра
зную кривую при ограничивающем воздействии лимитирующих факторов.
Главенствующее значение для естественного возобновления ценопопуляции имеет запас диаспор в почве, наличие подроста и занос зачатков. В ненарушенной лесной среде практически исключена смена лесного типа растительности на луговой, степной или болотный. Если же ширина окна превышает две и более высоты сомкнутого окружающего полога, то возникает нелесная среда, где преимущество заселения получают вначале травяной и моховой типы растительности, и только потом заселяются пионерные береза, осина, иногда сосна, при этом частично или навсегда исчезают многие лесные виды растений, животных и, вероятно, микроорганизмов.
Можно наблюдать следующие закономерности возникновения возобновления сообществ. Почвенный запас семян хорошо защищен от поражения микроскопическими грибковыми организмами, неспорообразующими бактериями, спорообразующими бактериями, актиномицетами, но недостаточно – от представителей лесной фауны, особенно видов, питающихся в основном семенами. Случайным является попадание семян в оптимальные условия прорастания, совпадающие с генетической наследственностью. При прорастании семени проросток сразу вступает в тесные взаимоотношения с микробиоценозом, эдафотопом и климатопом, т.е. любое растение образует с биоценозом консорцию. В этот период наблюдается массовая гибель растений из-за несовпадения консорциативных и неблагоприятных экотопических связей. Объем пространства, занимаемый проростками и ювенильными древесными растениями, невелик, и конкуренции за свет и почву между ними нет. Ресурсы среды у них могут отнять мхи, травы и взрослые древесные особи. Это предопределяет всегда недостаточное число возобновления на начальном этапе. Даже количество возобновления, равное 2,5 млн. шт./га, означает 40 см2 (5 x 8 см), приходящихся на одно растение. Следовательно, энергию роста (бонитет) на начальном этапе полностью определяют почвенное богатство и сопряженность с микробоценозом. В это время закладывается жизненная стратегия роста и развития (так называемые классы Крафта – деление деревьев в лесу на пять классов по росту и развитию: прегосподствующие, господствующие, умеренно развитые, заглушенные и подпологовые) каждой особи. По мере роста особь увеличивает занимаемое ею пространство до начала смыкания кронами с другими особями, после чего начинается закономерная межвидовая и видовая ценопопуляционная конкуренция между особями в биогруппе вначале за свет надземного пространства, позже присоединяется конкуренция корневых систем особей за почвенные ресурсы. По отношению к каждой особи этот процесс получил название «давление среды», приводящее к изреживанию в биогруппах и в насаждении в целом. Выжившие особи увеличивают консорциативные связи, вступая в тесные взаимоотношения со всем биоценозом, формируя собственную свиту растений, животных, грибов, микроорганизмов и изменяя внутреннюю среду биогеоценоза в выгодном для себя отношении. В метрическом исчислении это выражается двумя показателями – высотой и занимаемым воздушным и почвенным объемами, т. е. числом особей на 1 га. Чем беднее почва, тем больше необходимый занимаемый особью объем почвы, в связи, с чем площадь крон отстает от площади корневой системы, в результате исключается стопроцентная сомкнутость крон, а значит не полностью создается лесная среда. Особенно это видно в переходных типах растительности. При любом возрасте конкурентные отношения выражаются высотой и числом стволов, что позволяет сравнивать с высокой степенью корреляции эти показатели при разных бонитетах и возрастах.
Все модификаторы стремятся в течение первоначальных 20 лет занять ведущее место в верхнем пологе. Если коренные породы сосна, дуб, ель осуществляют это за счет долголетия, то короткоживущие пионерные береза и осина – путем интенсификации природных ресурсов в первое пятилетие, увеличивая прирост в высоту на относительно богатых почвах на один класс бонитета, по сравнению с коренными, и одновременно формируя тонкий ствол и небольшой объем кроны и сучьев. Рост в высоту эдификаторов сдерживают следующие лимитирующие факторы:
· годовая теплообеспеченность в лесной зоне снижается с юга на север на 0,51 о С на 1о широты, что приводит к падению производительности на 0,2 – 0,3 класса бонитета на каждый градус широты на богатых почвах;
· при достижении 7 – 10 метров многие виды вступают в период плодоношения, затрачивая ресурсы на семяношение;
· по достижении 15 м и выше увеличение кроны приводит к необходимости наращивать толщину ствола во избежание ветролома;
· при высоте 20 метров и выше требуется увеличение корневой массы для избежания ветровала.
В зоне смешанных лесов Русской равнины и подзоне южной тайги рост в высоту ограничивается для пихты, ели, липы, ольхи черной 30 м, осины, березы – 35 м, дуба – 40 м, лиственницы, сосны – 45 м. Для сравнения пихта кавказская достигает 50 – 60 м, иногда – 80 м, ель кавказская 50 иногда 65 м.
Если принять предельную ширину окна в 1,5 высоты полога и длину в две ширины окна, оптимальные размеры окна составят (исключая кавказские виды) соответственно 0,4; 0,6; 0,7; 0,9 га. Приняв ориентировочно для главных лесообразователей средний верхний предел возраста насаждений ели – 250 лет, сосны – 350 лет и дуба 400 лет, что дает вместе с окнами число парцелл, соответственно 26, 36, 41, а среднее число видов вместе с сопутствующими 3; несложным расчетом получаем минимальный размер постоянного экологического участка, равный для ели 0,6 ∙ 26 ∙ 3 = 47 га, сосны 0,9 ∙ 36 ∙ 3 = 97 га, дуба 0,7 ∙ 41 ∙ 3 = 86 га.
Колебания давления среды на каждую особь происходят, как правило, циклически. Так, по данным Н. Я. Саликова, полноты древостоев колеблются от критических самоизреживающизся 0,67 до максимальных предельно плотных 1,33. Колебания давления среды связаны с увеличением объема пространства дерева во время роста. С учетом микроколебаний, богатства почвы, разной энергии роста деревьев, некомпактного (по сравнению с компактным равносторонним треугольным расположением) размещения по площади, выпадения отставших в росте, что освобождает ресурсы для оставшихся, давление среды всегда неодинаковое для отдельно рассматриваемых особей. До определенного возраста (примерно 100 лет) отпад положительно влияет на рост и развитие древостоя. Это этап динамических изменений – сукцессия, происходящая непрерывно, но с большими изменениями среды в первоначальном периоде от стадии окна и постепенным замедлением изменений, примерно, до 100 лет, после чего насаждение входит в этап гомеостаза. Ю. Одум (1980 г.) под экологической сукцессией понимает вообще весь процесс развития экосистемы. Более конкретное определение дает этому явлению Н. Ф. Реймерс (1990 г.): «Сукцессия – последовательная смена биогеоценозов, приемственно возникающая на одной и той же территории (биотопе) под влиянием природных факторов (в том числе и внутренних противоречий самих биоценозов) или воздействия человека». Изменения в сообществе в результате сукцессии носят закономерный характер и обусловлены взаимодействием организмов между собой и с окружающей абиотической средой.
Другие рефераты на тему «Экология и охрана природы»:
Поиск рефератов
Последние рефераты раздела
- Влияние Чекмагушевского молочного завода на загрязнение вод реки Чебекей
- Влияние антропогенного фактора на загрязнение реки Ляля
- Киотский протокол - как механизм регулирования глобальных экологических проблем на международном уровне
- Лицензирование природопользования, деятельности в области охраны окружающей среды и обеспечения экологической безопасности
- Мировые тенденции развития ядерной технологии
- Негативные изменения состояния водного бассейна крупного города под влиянием деятельности человека
- Общественная экологическая экспертиза и экологический контроль