Катаболизм и стресс у растений
Старение организмов приводит к постепенному уменьшению величины отношения анаболизм/катаболизм. В различных органах растений степень усиления катаболизма с увеличением общего возраста организма может отличаться в зависимости от собственного возраста, например, у метамерных органов (листьев). Усиление катаболизма старых листьев способствует «перекачке» образующихся мономеров (и продуктов их прев
ращения в легко транспортируемые формы) и реутилизации органических соединений в более молодых или репродуктивных органах. Среди неспецифических ультраструктурных изменений, вызванных стрессом и старением, можно отметить изменение тонкой гранулярной структуры ядра, уменьшение числа полисом и диктиосом, набухание митохондрий и хлоропластов, уменьшение в хлоропластах числа тилакоидов, перестройку цитоскелета [3, стр.157].
В изучении процессов деградации у растений важную роль играет выбор объекта исследований. Например, сильная интенсификация катаболизма биополимеров и липидов наблюдается у прорастающих семян; в органах растений с быстрым темпом старения (например, у лепестков некоторых видов растений); у затемненных листьев; в участках корней, заканчивающих рост растяжением [4, стр.197]; в тканях и клетках, разрушающихся в результате механического повреждения, подвергающихся действию бактерий, грибов, вирусов, эколого - климатических или антропогенных стрессоров. Во многих случаях используются изолированные протопласты, культуры клеток или каллусных тканей растений.
Анализ событий, совершающихся в клетках с момента начала действия того или иного стрессора, заставляет считать, что имеется небольшое число наиболее лабильных (чувствительных) компонентов клеточных структур и звеньев метаболизма, которые выполняют роль пускового механизма, вызывающего последующие глобальные нарушения, проявляющиеся в виде неспецифических изменений в обмене веществ.
По всей вероятности, функцию спускового крючка, на которых "нажимают" разнообразные стрессоры и который приводит в действие сложный механизм ответа, в большинстве случаев выполняют мембранные системы.
Мембраны - это "ахиллесова пята" клеток, в первую очередь повреждающаяся под влиянием экстремальных факторов. Совершенно очевидно, что во многих случаях, например, при изменении температуры, роль сенсорного компонента мембран играют липиды, от состава и соотношения которых зависит степень вязкости мембран, определяющая структуру и функции мембранных белков.
Изменение состояния мембран приводит к включению процесса деградации липидов, от которого зависит реализация программы формирования стресса растительных клеток[4, стр.197].
3. БИОГЕННЫЙ СТРЕСС И КАТАБОЛИЗМ
В предыдущем разделе уже затрагивался вопрос о влиянии патогенов на образование углеводных элиситоров и белковый обмен. В последние годы появляется все больше информации о метаболическом взаимодействии патогенных организмов - бактерий и грибов - и тканей различных органов растения-хозяина. Первый этап этого взаимодействия - конформационное узнавание поверхностей органов и клеток патогена и хозяина. Второй - экскреция клетками патогенных бактерий и грибов ферментов, гидролизующих биополимеры и липиды покровных тканей растения-хозяина, разрыхляющих их и обеспечивающих более интенсивное проникновение патогена в ткани хозяина и обильное питание, необходимое для развития патогена. Третий - образование в ходе деградации биополимеров и липидов различных физиологически активных промежуточных продуктов - элиситоров, выполняющих роль сигнальных веществ - стимуляторов ответной реакции клеток хозяина. Элиситоры непосредственно или с помощью посредников влияют на генетический аппарат клеток хозяина, вызывая (четвертый этап) синтез веществ, способствующих или повышению устойчивости к патогену, или вызывающих сверхчувствительность и гибель клеток, но тем самым создающих механический барьер, препятствующий распространению инфекции по тканям растения (рис. 2). Промежуточные продукты катаболизма клеток хозяина могут выступать и в роли эффекторов метаболизма и развития патогена [5, стр.44].
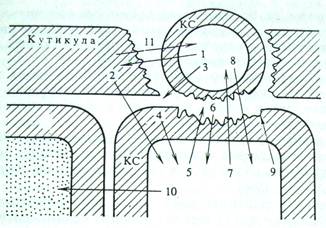
Рис. 2. Схема взаимодействия клетки патогена с растением- хозяином
1 — кутиназа; 2 — продукты деградации компонентов кутикулы (возможно, обладающие сигнальными свойствами); 3 — β-глюканаза и другие гликозилазы, экскретируемые патогеном; 4 — элиситоры — фрагменты клеточной стенки (КС) хозяина; 5 — хитиназы и другие гликозилазы, действующие разрушающе на КС патогена; 6 — элиситоры — фрагменты КС патогена; 7 — фитоалексины — ингибиторы протеиназ, кутиназ, гликозилиз и других ферментов патогена; 8 — токсические вещества патогена; 9 — укрепление КС хозяина за счет активации пероксидаз и усиления синтеза лигнина. Отложение оксипролиновых белков, лектинов; 10 — вещества — индукторы сверхчувствительности и некроза соседних клеток; 11 — продукты деградации кутина, действующие на клетку патогена
Наружным покровом растений является кутикула, состоящая главным образом из гетерополимера кутина, погруженного в воск. Обнаружено более 20 мономеров, из которых состоит кутин: различной длины насыщенные и ненасыщенные жирные кислоты и спирты, в том числе гидроксилированные и эпоксидированные, дикарбоксиловые кислоты и т.д. В кутине большинство первичных спиртовых групп участвует в образовании эфирных связей, так же как часть вторичных спиртовых групп, обеспечивающих сшивки между цепями и точки ветвления в полимере. Другой "барьерный" полимер - суберин, состоит, по-видимому, из фенольных и алифатических доменов, первые из которых близки по своему составу к лигнину, а вторые - к кутину. Отличия алифатического домена в том, что свободные жирные кислоты являются главным компонентом субериновых восков, в то время как в кутине их очень мало. Кроме того, в суберине присутствуют главным образом С22и С24 жирные спирты, в то время как в кутине – С26 и С 28 [5, стр.44].
Оказалось, что многие патогенные грибы могут выделять ферменты, гидролизующие кутин и суберин. При гидролизе лигниноподобной фракции суберина образовывались кумаровая и феруловая кислоты, причем большая часть фракции оставалась негидролизованной. Продуктами кутиназной реакции были различные оксигенированные жирные кислоты и спирты [5, стр.45].
По всей вероятности, в спорах грибов кутиназа содержится в очень небольших количествах, и при контакте с кутикулой растений гидролизу подвергается лишь малая часть кутина. Однако образующиеся активные сигнальные молекулы - 10,16-дигидрокси-С16- и 9,10,18-тригидрокси-С18-кислоты - транспортируются в прорастающую спору и индуцируют образование больших количеств дополнительной кутиназы, начинающей интенсивное разложение кутина и облегчающей инфицирование растения. Было обнаружено, что лаг-период появления кутиназной м-РНК после начала действия ди- и триоксикислот составляет всего 15 мин, а появления кутиназы - в два раза больший [5, стр.45]. Ингибирование кутиназы с помощью химических препаратов или антител предотвращало инфекцию.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики