Отдел красные водоросли
На гаметофитах половые органы возникают на трихобластах — веточках, имеющих моносифоновое строение, т. е. сложенных из одного ряда клеток и не окруженных перицентральными клетками. Плодущая часть мужского трихобласта становится полисифоновой в результате отчленения продольными перегородками перицентральных клеток. Они становятся материнскими клетками спермациев. Спермации отпочковываются послед
овательно: одновременно каждая материнская клетка спермация может нести 2—6 спермациев различного возраста. Спермации соединены с соответствующими материнскими клетками посредством пор. Заново отчленившийся спермации содержит митохондрии; ядро располагается в его базальной части — ближе к материнской клетке, дикти-осомы и вакуоли вначале отсутствуют. По мере отпочковывания от материнской клетки последующих спермациев содержимое ранее отчленившегося спермация претерпевает изменения: ядро мигрирует и занимает апикальное положение; в базальной части спермация возникают цистерны шероховатого эндоплазматического ретикулума, которые сливаются с образованием двух «спермациальных» вакуолей. Сначала эти вакуоли мелки и имеют гранулярное содержимое, позднее превращающееся в фибриллярный материал. После формирования вакуолей в спермации появляются диктиосомы, везикулы которых секретируют материал стенки спермация. Образующаяся вокруг спермация слизистая стенка содействует разрыву септальной пробки, что облегчает освобождение спермация. При этом спермациальные вакуоли выделяются путем экзоцитоза. Кроме того, по мере отчленения от материнской клетки последующих спермациев и их увеличения в объеме они оказывают давление на более старые спермации сбоку и снизу, выталкивая их наружу. В результате этого спермации энергично извергаются, а в пространство, остающееся на месте вытолкнутого спермация, может врастать добавочный спермации. Дифференцировка спермация, включающая развитие «спермациальной» вакуоли и ее экзоцитоз, а также образование диктиосомами слизистой стенки спермация, возможно, имеющей значение для прилипания его к трихогине, наблюдалась и у других церамиальных, например у Ptilota plumosa.
Плодущий сегмент женского трихобласта формирует пять перицентральных клеток: две на нижней стороне, по одной с боков и, наконец, одну перицентральную клетку сверху — она и является поддерживающей — несущей клеткой, на которой развивается четырехклеточная карпогонная ветрь. Карпогон одноядерный, трихогина безъядерна. После оплодотворения от несущей клетки отчленяется ауксилярная клетка, диплоидное ядро оплодотворенного брюшка карпогона делится митотически и одно из дочерних ядер переходит в ауксилярную клетку при непосредственном слиянии карпогона и ауксилярной клетки. Собственное гаплоидное ядро ауксилярной клетки дегенерирует, а диплоидное делится и затем от ауксилярной клетки наверх и кнаружи отчленяется первая гонимобластная клетка, из которой развиваются гонимобластные нити: их конечные клетки представляют собой карпоспорангии. Из боковых перицентральных клеток плодущего и базального сегментов трихобласта еще до оплодотворения развиваются нити, которые образуют вокруг прокарпия и развивающихся карпоспор псевдопаренхиматозную оболочку с отверстием на вершине. Через него по созревании выпадают карпоспоры. Диплоидные карпоспоры прорастают в диплоидные тетраспорофиты, морфологически сходные с гаметофитами, но несущие органы бесполого размножения — тетраспорангии. Тетраспорангии возникают на ветвях полисифонового строения таким образом, что плодущая перицентральная клетка сначала отчленяет налево и направо две покровные клетки, а затем делится косо на нижнюю более мелкую и верхнюю более крупную клетку, которая и становится спорангием. После мейоза в спорангии формируются четыре тетраэдрически расположенные тетраспоры. Тетраспоры прорастают в гаплоидные гаметофиты, несущие половые органы.
Среди церамиальных, как и у артинальных, в последние годы описаны случай аномального сочетания репродуктивных органов. Так, у Polysiphonia harlandii 14% образцов, собранных в мае 1984 г. в литоральной зоне Гонконга, аесли одновременно а тетраспоры, и цистокарпии.
Одновременное развитие цистокарпиев и тстраспорангиев на некоторых талломах отмечено и для Р. тасгосагра. Ранее у другой церамиальной водоросли Antithamnhn tenuissimum были обнаружены некоторые мужские растения, несущие тетраспорангии. При спаривании этих мужских растений с нормальными женскими получены карпоспоры, развившиеся в нормальные тетраспорофиты, Последние, в свою очередь, дали начало нормальным мужским и женским растениям и некоторым мужским растениям, несущим тетраспоры.
Заслуживает внимания род гриффитзия, виды которого благодаря простоте строения таллома и очень крупным клеткам стали объектами интересных экспериментов. Некоторые из них приводятся ниже.
Род гриффитзия отличается очень крупными, не одетыми корой многоядерными клетками, легко различимыми невооруженным глазом. Таллом вертикальный, разветвленный, прикрепленный к субстрату ризоидами. Весь таллом заключен в обильную слизь. Короткие веточки, несущие репродуктивные органы, возникают от верхушек более молодых клеток, то же положение занимают обильно разветвленные волоски. Спорангии развиваются на специальных боковых веточках, которые образуют мутовки у верхушек сегментов. Каждая фертильная боковая ветвь состоит из одной клетки, несущей несколько спорангиев, которые защищены выростами, возникающими от той же клетки или от родительского сегмента. Женские плодущие веточки состоят из нескольких многоядерных клеток и берут начало от верхушки крупных осевых клеток. Из самой нижней клетки веточки позднее возникает обвертка цистокарпия, а из субтерминальной клетки—прокарпий. Она отчленяет три перицентральные клетки: те из них, которые отчленяются направо и налево, развиваются в двухклеточные веточки; их базальная клетка несет карпогонную ветвь.
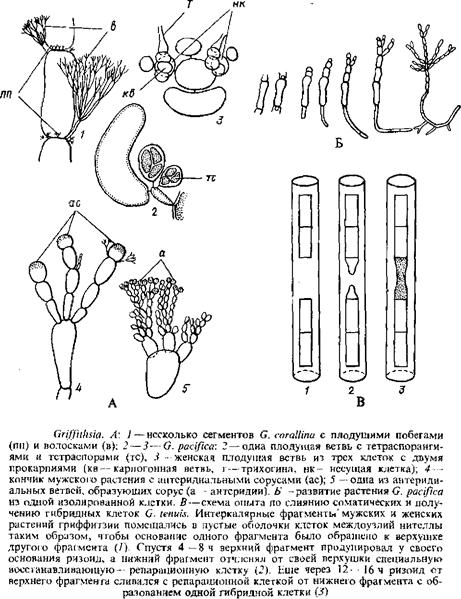
Осуществляется непосредственное слияние ауксилярной клетки и карпогона: специальная соединительная клетка не образуется. Ауксилярная клетка продуцирует несколько зачатков гонимобластов, которые последовательно развиваются в обильно разветвленные гонимобласты; большинство их клеток формирует одноядерные карпоспоры. Наблюдается заметное увеличение пор и слияние между клетками плодущей веточки, ауксилярной клетки с несущей и др. Несколько одевающих цистокарпии нитей имеют только две клетки. Антеридиальные сорусы состоят из плотно разветвленных веточек, сформированных главным образом одноядерными клетками. Они образуют «шапочки» на терминальных сегментах таллома или окружают перетяжки между более молодыми клетками.
Хотя у G. pacifica вертикальные талломы и ризоидальные нити нарастают только за счет деления их апикальных клеток, любая изолированная интеркалярная клетка таллома способна регенерировать весь таллом. Пусковым механизмом этого процесса служит удаление примыкающих клеток. Так, в опытах по регенерации для изолирования отдельной клетки соседние с ней клетки отрезались. Такая живая клетка помещалась на питательную среду и при определенных внешних условиях уже в течение первых суток отчленяла от своей верхушки клетку будущего вертикального побега, а у своего основания—ризоидальную клетку. Дальнейшее удлинение побега осуществляется за счет деления апикальной клетки, когда нить достигает длины 4—5 клеток, возникают первые боковые ветви. На 5-й день изолированная из побега клетка уже образует растение, различимое невооруженным глазом.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики