Свободные аминокислоты нервной системы
Активность систем транспорта аминокислот, так же как и состав их пула, изменяется в процессе развития мозга. Аминокислоты проникают в мозг молодых животных быстрее и достигают более высоких концентраций, чем у взрослых.
В литературе отсутствуют сообщения о болезнях, вызванных нарушением транспорта аминокислот в мозг, вероятно, потому, что они летальны. Даже дефекты транспорта аминокислот в
другие ткани ведут к заболеваниям, имеющим неврологические последствия.
Наряду с неопасным для жизни синдромом Хартнупа, вызванным дефектом транспорта триптофана в малый кишечник и почки и схожим клинически с пеллагрой, известен ряд недугов с тяжелыми неврологическими последствиями, также обусловленных дефицитом поступления аминокислот. Среди них - цистиноз - нарушение транспорта цистина в клетки, особенно почек; цисти-ноз сопровождается фотофобией и повреждением глаз. Тяжелым, нередко летальным заболеванием, связанным с транспортом аминокислот в кишечник, является окулоцеребральный синдром. Он сопровождается глаукомой, катарактой, слепотой. Перечень этих болезней, вызванных нарушением транспорта триптофана, метионина, нейтральных и других аминокислот в кишечнике и других органах, довольно велик, причем все они косвенно затрагивают уровень аминокислот в мозге и имеют поэтому неврологические проявления.
2. Метаболизм дикарбоновых аминокислот и глутамина
Более 2/3 аминоазота аминокислот приходится на долю глутамата и его производных; эти аминокислоты доминируют в количественном отношении в мозге всех изученных видов животных. В спинном мозге наблюдается аналогичная картина, а периферическая нервная система содержит значительно меньше глутамата, глутамина, N-ацетиласпартата, чем головной мозг, а ГАМК почти отсутствует в периферических нервах позвоночных. При высоком уровне этих аминокислот в головном мозге метаболизм их также чрезвычайно быстрый.
3. Глутамат и аспартат
Особенностью метаболизма глутамата в нервной ткани является его тесная связь с интенсивно функционирующим в этом органе циклом трикарбоновых кислот, что и позволяет считать его промежуточным продуктом энергетического метаболизма. Так, уже через 30 мин после инъекции меченой глюкозы более 70% радиоактивности растворимой фракции приходится на долю глутамата и его производных. Этому способствует чрезвычайно быстрое взаимопревращение глутамата и а-кетоглутарата в ЦНС. Высокий процент включения радиоактивности из глюкозы в аминокислоты мозга явился основанием для предположения, что утилизация глюкозы в этом органе в значительной степени происходит через биосинтез и окисление аминокислот.
Непосредственным предшественником для синтеза глутамата в мозге является а-кетоглутаровая кислота, которая может превращаться в глутамат или путем прямого восстановительного аминирования с участием глутаматдегидрогеназы, или путем переаминирования.
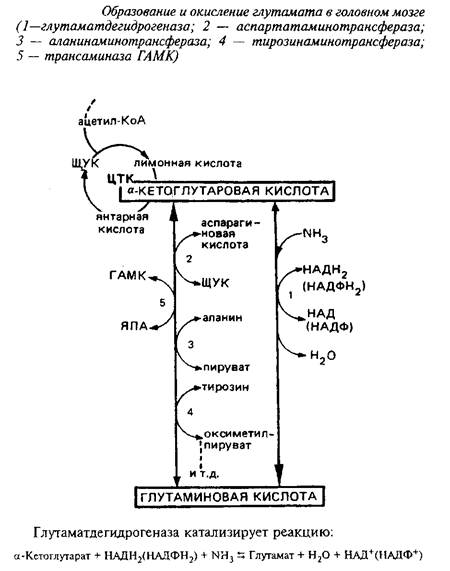
Энзим менее активен в мозге, чем в печени, присутствует в митохондриях, требует в качестве кофакторов пиридиннуклеотидов и активируется АДФ. Км этого энзима для аммония близок к 8 мМ. Реакция обратима, однако равновесие сильно сдвинуто в сторону прямой реакции, т.е. синтеза глутаминовой кислоты.
Таким образом, в головном мозге глутаматдегидрогеназная реакция участвует не столько в окислении глутамата, сколько в синтезе его из а-кетоглутаровой кислоты, обеспечивая тем самым непрерывное превращение свободного аммиака в аминоазот аминокислот. Основной же путь окисления глутамата в мозге - через переаминирование.
В митохондриях мозга 90% глутамата подвергается переаминированию с образованием аспартата. Фермент, катализирующий переаминирование глутамата с щавелевоуксусной кислотой, - аспартатаминотрансфераза является наиболее мощной трансаминазой головного мозга. Выделены два изоэнзима аспартатаминотрансферазы, локализованных в митохондриях и цитоплазме. Функциональная роль их различна. Митохондриальный фермент связан в основном с функционированием ЦТК, цитоплазматический определяет интенсивность глюконеогенеза.
Как уже отмечалось, путь метаболизма глутамата через переаминирование намного активнее дегидрогеназного. В регуляции соотношения между этими двумя путями, конкурирующими за один субстрат, важная роль принадлежит макроэргическим соединениям. В интактных митохондриях энзим взаимодействует по преимуществу с НАДФ+ и интенсивность реакции пропорциональна отношению НАДФ+/НАЦФН2. Макроэргические соединения способствуют превращению НДЦФ+ в НАДФН2 и тем самым подавляют дезаминирование глутамата. Наоборот, трансаминазный путь требует расходования макроэргических соединений. Поэтому выбор между этими двумя реакциями определяется энергетическими возможностями митохондрий.
При нормальном функционировании ЦТК дегидрогеназный путь окисления глутамата подавлен, а трансаминазный активно протекает. В результате уменьшения количества макроэргических соединений, например при добавлении к митохондриям разобщителя окислительного фосфорилирования 2,4-динитро-фенола, подавляется трансаминазный путь при одновременном резком усилении дегидрогеназного пути окисления глутамата.
Взаимопревращение а-кетоглутарата и глутамата происходит чрезвычайно быстро. В мозге был идентифицирован метаболический путь такого взаимопревращения, получивший название аспартат-малатного шунта, служащего для транспорта восстановительных эквивалентов из цитозоля в митохондрии.
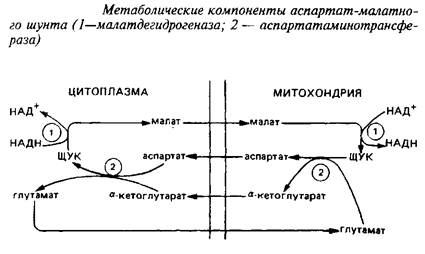
Уже упоминалось, что различные органеллы клеток мозга могут индивидуально контролировать уровни аминокислот, накапливая их против градиента концентрации. Примером этого могут служить изолированные из ЦНС митохондрии, которые быстро поглощают глутамат и малат, освобождая соответствующие количества аспартата и а-кетоглутарата. Это означает, что ток аспартата через митохондриальную мембрану связан с током глутамата в обратном направлении; также реципрокно связаны ток малата и а-кетоглутарата. Энзимы, катализирующие отдельные реакции малат-аспартатного шунта, превалируют в тканях ЦНС. В нейронах малат-аспартатный шунт является преобладающим механизмом переноса восстановительных эквивалентов в митохондрии.
Таким образом, глутаминовая кислота выполняет чрезвычайно важную функцию в энергетическом обеспечении головного мозга, которая заключается в поддержании метаболитов ЦТК на определенном и довольно высоком уровне, а также в снабжении митохондриальных синтетических процессов восстановительными эквивалентами.
Большое значение имеет образование аммиака из глутамата. В головном мозге обнаружены многочисленные аминотрансферазы основных, кислых, нейтральных и ароматических аминокислот. При участии этих ферментов аминогруппы различных аминокислот переносятся в конечном счете на глутамино-вую кислоту. Последняя переаминируется с ЩУК при участии аспартатами-нотрансферазы с образованием аспартата. Образование аммиака из аспартата происходит различным образом в митохондриях и цитоплазме. В митохондриях этот процесс связан с аминированием дезаминоформ НАД+ и включает в себя три ферментативных реакции.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики