Низшие хордовые и их филогенетические связи с другими типами животных
Полухордовые в качестве основного органа движения стали использовать предротовой отдел тела: хобот кишечнодышащих или головной щит крыложаберных. Обычно полухордовых считают близкими предками хордовых, о чем говорит и само название этой группы.
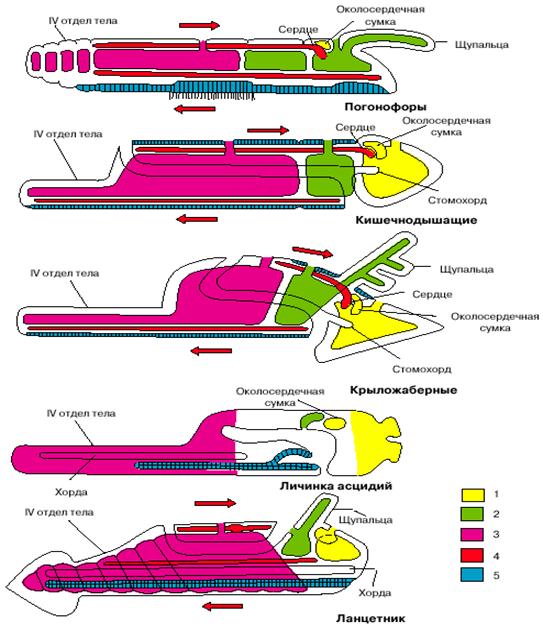
Рис. 5. Сравнение планов строения различных групп вторичнорот
ых (личинка асцидий и ланцетник изображены в перевернутом положении). 1 - целомы первой пары, 2 - целомы второй пары, 3 - целомы третьей пары, 4 - кровеносная система, 5 - нервная система. Стрелки показывают направление тока крови.
В действительности эта группа не слишком близка к хордовым, в частности потому, что имеет два нервных тяжа, только один из которых может быть гомологизирован с нервной трубкой хордовых. У них имеется слепой вырост кишечника - стомохорд, который сформировался как опорная структура хобота и не имеет прямого отношения к хорде хордовых животных.
Современные погонофоры - сидячие организмы, обитающие внутри выделяемых ими трубок. Во взрослом состоянии они лишены кишечника и питаются за счет симбиотических хемосинтезирующих бактерий, окисляющих сероводород или метан. Их предки, вероятно, питались этими бактериями, собирая их из придонного слоя воды с помощью щупалец.
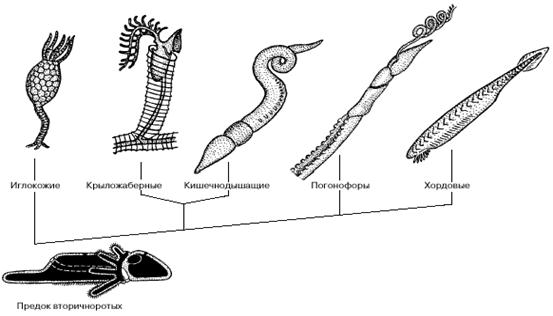
Рис. 6. Филогенетические отношения между вторичноротыми животными.
Впоследствии они стали культивировать бактерий в клетках кишечника. Личинки одной из групп современных погонофор - вестиментифер - на ранних стадиях развития собирают бактерий с поверхности грунта, а затем начинают их культивировать внутри видоизмененного кишечника, после чего рот и анус у них редуцируются. Особенности эмбрионального развития и строения нервной системы позволяют рассматривать погонофор как самую близкую к хордовым группу вторичноротых.
Предки хордовых также первоначально собирали пищевые частицы с помощью околоротовых щупалец, которые до сих пор сохранились у ланцетников. Одной из особенностей эволюции хордовых явилось использование самого заднего, четвертого отдела тела как органа активного движения. Этот отдел имеется у всех вторичноротых. Иглокожие и крыложаберные используют его как прикрепительный стебелек. У кишечнодышащих этот отдел функционирует только у молодых особей: с его помощью они закапываются в толщу грунта. У погонофор этот отдел активно функционирует всю жизнь: он вооружен щетинками, торчит из заднего конца трубки и благодаря его движениям погонофоры заглубляют задний конец трубки в толщу грунта по мере своего роста. Хордовые - единственная группа вторичноротых, использующая этот отдел как орган активного плавания. Именно в связи с активной двигательной функцией этого органа в нем происходит полная сегментация мускулатуры и развивается хорда (у личинок оболочников хорда локализована именно в хвосте. Другой важный момент эволюции хордовых - это инверсия сторон тела, то есть произошедший в филогенетическом развитии хордовых переворот с морфологически брюшной на морфологически спинную сторону. Такой переворот мог совершиться при переходе от улавливания с помощью щупальцевого аппарата взвешенных в толще воды пищевых частиц, как это делают, например, современные крыложаберные, к их сбору из поверхностного слоя грунта, как это делают современные ланцетники.
Заметим, что между личинками оболочников, с одной стороны, и бесчерепными и позвоночными, с другой, есть одно важное различие. Если внимательно посмотреть на две нижние фигуры на рис. 5, изображающие личинку асцидии и ланцетника, то нетрудно заметить, что у первых рот располагается на той же стороне, что и нервная трубка (последняя даже открывается в глотку особым отверстием), то есть на исходно брюшной стороне, а у ланцетника (так же, как у позвоночных) - на противоположной стороне тела. Положение рта у личинок асцидий - первичное, оно совпадает с тем, что имеет место у других вторичноротых (см. рис. 4). Таким образом, личинки асцидий по своей организации более примитивны, чем остальные хордовые, и отражают организацию подвижных предков хордовых до их переворота.
У бесчерепных и позвоночных рот располагается на противоположной стороне, гомологичной спинной стороне тела прочих вторичноротых (см. рис. 5). Такое положение рта не может быть первичным и возникло уже после переворота. Удивительно, но в индивидуальном развитии бесчерепных сохранились следы постепенного перемещения рта со спинной стороны на брюшную по левой стороне тела. А один из представителей бесчерепных (Assymetron) так и застыл на стадии, когда рот остался на левой стороне тела. Все это отголоски постепенного перемещения рта со старой морфологически брюшной стороны на новую физиологически брюшную сторону.
6. ОСНОВНЫЕ ГИПОТЕЗЫ ПРОИСХОЖДЕНИЯ ХОРДОВЫХ
Одна из первых концепций, связывающих план строения хордовых с планами строения других групп беспозвоночных, была разработана выдающимся французским сравнительным анатомом Жоффруа Сент-Илером еще в первой половине XIX века. Он считал, что членистые животные (кольчатые черви и членистоногие) могут рассматриваться как перевернутые позвоночные. "Взгляните на рака, опрокинутого на спину, говорил он, - и вы увидите, что различные системы его расположены совершенно так же, как у высших позвоночных животных". Именно эта концепция была в центре публичного спора между Жоффруа Сент-Илером и Жоржем Кювье, который состоялся в Париже в 1830 году в дни июльской революции, свергнувшей Бурбонов. Как известно, в том знаменитом споре победил Жорж Кювье. Однако к идее сравнения хордовых (и позвоночных) и перевернутых кольчатых червей или членистоногих зоологи возвращались неоднократно на протяжении всего XIX века.
В начале нынешнего столетия были разработаны гипотезы происхождения хордовых от кишечнодышащих - обитающих в толще грунта морских червеобразных организмов. У этих животных (как и у хордовых) имеются жаберные щели, а в передней части тела - опорный вырост кишечника (стомохорд), который сравнивали с хордой. Однако кишечнодышащие резко отличались от хордовых обратным направлением тока крови, расположением сердца, строением нервной системы и другими важными особенностями организации. Делались попытки выводить хордовых от иглокожих, немертин, моллюсков и других групп беспозвоночных, однако ни одна из них не имела успеха.
Своеобразной реакцией на неудачи зоологов решить проблему происхождения хордовых традиционными способами явилась разработка так называемых неотенических гипотез, выводящих хордовых от личинок асцидий. Взрослые асцидии - прикрепленные морские животные, фильтрующие воду через объемистый жаберный мешок и улавливающие взвешенные в воде мельчайшие органические частицы. Они не имеют ни хорды, ни нервной трубки и совсем не похожи на хордовых животных. Но, как давно известно (а впервые это показал еще в конце прошлого века выдающийся русский биолог А.О. Ковалевский), личинки асцидий имеют и хорду, и нервную трубку, которые утрачиваются при метаморфозе. На основании этого многие биологи (Берилл, Уитер, Боун и др.) предполагают, что хордовые произошли от ставших половозрелыми личинок асцидий. Правда, в этом случае остается непонятным, откуда взялись сами асцидии. Да и трудно представить, чтобы хорда и нервная трубка возникли у живущих 1 - 2 дня и не питающихся личинок. Более вероятно, что эти особенности - наследие свободноживущих предков асцидий, об организации которых неотенические гипотезы ничего не говорят. Ни одна из рассмотренных гипотез в настоящее время не может считаться истинной, но в каждой из них содержатся отдельные элементы, которые могут оказаться полезными при реконструкции путей происхождения хордовых.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики