Особенности строения нервной системы
Липиды центральной и периферической нервной системы
Фосфолипиды, фосфатиды, сложные липиды, отличительным признаком которых является присутствие в молекулах остатка фосфорной кислоты. В состав Ф. входят также глицерин (или аминоспирт сфингозин), жирные кислоты, альдегиды и азотистые соединения (холин, этаноламин, серин). Важнейшие представители Ф. – глицерофосфатиды [фосфатидилхолин (лецити
н), фосфатидилэтаноламин (устаревшее название – кефалин), фосфатидилсерин, фосфатидилинозит, кардиолипин] и фосфосфинголипиды – сфингомиелины. Каждый класс Ф. объединяет множество однотипных молекул, содержащих различные жирные кислоты или альдегиды. При этом ненасыщенные жирные кислоты преимущественно находятся при 2‑м углеродном атоме молекулы глицерина (формулы см. в ст. Липиды).
Ф. широко распространены в природе. В качестве основных структурных компонентов они входят в состав клеточных мембран животных, растений и микроорганизмов, определяя их строение и проницаемость, а также активность ряда локализованных в мембранах ферментов. С белками Ф. образуют липопротеиновые комплексы. Различным биологическим мембранам присущ определённый состав Ф. Так, кардиолипин – специфический митохондриальный Ф.; сфингомиелин присутствует в основном в плазматических мембранах. В мембранах микроорганизмов всегда содержится фосфатидилглицерин и редко лецитин (в отличие от клеток животных).
Состав Ф. некоторых органов изменяется при старении и ряде патологических состояний организма (атеросклероз, злокачественные новообразования).
Для разделения и установления строения Ф. используют различные виды хроматографии, химический и ферментативный (с помощью фосфолипаз) гидролиз, физические методы исследования (масс-спектрометрия, ИК-спектрометрия, ядерный магнитный резонанс и др.).
Помимо Ф., известны также фосфонолипиды, в которых атом фосфора связан с азотистым основанием (холином и этаноламином) ковалентной Р-С-связью. Эти соединения обнаружены у ряда моллюсков и бактерий.
Миелин и его роль в нервной системе
Миелин – сложная смесь белков и фосфолипидов, образующая внутреннюю часть оболочки некоторых типов нервных волокон
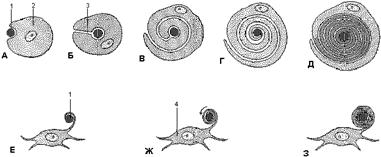
А-Д – последовательные стадии миэлинирования периферических нервных волокон или аксонов нейрилемной или Шванновской клеткой. Аксон сначала образует вмятину на клетка (А), а затем Шванновская клетка начинает окружать аксон, в то время как мезаксон (место инвагинации) удлинняется (Б). Затем Шванновская клетка начинает обвивать аксон (В-Г). Цитоплазма между слоями клеточных мембран постепенно конденсируется, Сохраняется цитоплазма вне слдоев миэлина и аксона (Д). Е-З – последовательные стадии миэлинирования нервных волокон в центральной нервной системе с помощью олигодендроцитов. Отросток нейроглиальной клетки обвивается вокруг аксона (Е), а добавляющиеся слои цитоплазмы движутся по направлению к телу клетки (Ж и З) 1 – аксон, 2 – нейрилемная (Шванновская клетка), 3 – мезаксон, 4 – олигодендроцит
Нуклеиновые кислоты мозга
Нуклеиновые кислоты – репликация ДНК в нейронах отсутствует, работает система репарации ДНК, в мозге экспрессируется несколько десятков тысяч уникальных генов, из которых не менее половины экспрессируется только в головном мозге – это говорит о высокой скорости транскрипции РНК, широко распространен альтернативный сплайсинг и интенсивное образование белка. Синтеза пиримидиновых нуклеотидов не происходит, т. к. нет карбамоилфосфатсинтетазы, для синтеза пуринов все есть. Содержание циклических нуклеотидов очень высокое, т. к. они участвуют в синаптической передаче нервного импульса.
Углеводы и их обмен в нервной системе
Спецификой углеводного обмена нервной системы является исключительная роль глюкозы для мозга. Глюкоза является основным источником энергии, так как клетки мозга не содержат ферментов для метаболизма липидов и других источников энергии.
Некоторая часть нейроглии способна запасать небольшое количество гликогена. Нейроны этой способностью не обладают.
Важной особенностью является повышенная доля обмена ди- и трикарбоновых кислот между матриксом митохондрий и цитозолем. Трикарбоновые кислоты с этом случае включаются в синтез нейромедиаторов.
Особая роль аэробных превращений глюкозы в энергетике мозга. Доля и роль гликолиза. Дополнительные источники энергии мозга. Основные системы, потребляющие энергию в мозгу (поддержание потенциала клеточных мембран, синтез белков, в особенности тубулярных, и др.).
Энергетический метаболизм мозга
Особенности обмена в нервной ткани:
1) много липидов, мало углеводов, нет их резерва
2) высокий обмен дикарбоновых кислот
3) глюкоза – основной источник энергии
4) мало гликогена, поэтому мозг зависит от поступления глюкозы с кровью
5) интенсивный дыхательный обмен
6) кислород используется постоянно и уровень не меняется
7) обменные процессы носят обособленный характер благодаря гематоэнцефалическому барьеру, высокая чувствительность к гипоксии и гипогликемии.
Гипоксия и окислительный стресс
Известно, что гипоксические, нейродегенеративные и возрастные нарушения в мозге характеризуются одними и теми же особенностями, в частности, накоплением активных форм кислорода (АФК). Выяснены тонкие молекулярные механизмы окислительного стресса в мозге и показана защитная функция природных антиоксидантов против апоптоза нейрональных клеток. Обнаружено, что Na/K‑АТФаза нейрональных мембран является мишенью для окислительного стресса. Установлен молекулярный механизм повреждения Na‑насоса, заключающийся в окислении сульфгидрильных групп и нарушении межсубъединичных взаимодействий в олигомерном комплексе фермента. Прослежена взаимосвязь между экзайтотоксическим действием глутамата и его агонистов на нейроны коры головного мозга и изменением активности Na/K‑АТФазы.
Систематические исследования антиоксидантной активности природного нейропептида карнозина показали его высокую эффективность по защите нейронов как в условиях in vitro (индивидуальные реакции повреждения макромолекул, суспензии изолированных нейронов или срезов мозга в условиях свободнорадикальной атаки), так и in vivo – на различных моделях экспериментальной ишемии мозга и сердца, гипобарической гипоксии, и т.д.
Установлено, что карнозин является важным природным фактором системы антиоксидантной защиты мозга в условиях окислительного стресса. Профессору А.А. Болдыреву и его ученикам принадлежит приоритет на практическое использование природного дипептида карнозина в качестве профилактического и терапевтического средства (имеются авторские свидетельства и патенты).
Нейроспецифичные белки
S‑100 является специфическим белком астроцитарной глии, способным связывать кальций. Свое название белок получил благодаря свойству оставаться в растворенном состоянии в насыщенном растворе сульфата аммония. Семейство белков S‑100 состоит из 17 тканеспецифичных мономеров, два из которых: α и β образуют гомо- и гетеродимеры, присутствующие в высокой концентрации в клетках нервной системы. S‑100 (ββ) присутствует в высоких концентрациях в глиальных и шванновских клетках, гетеродимер S‑100 (αβ) находится в глиальных клетках, гомодимер S‑100 (αα) – в поперечнополосатых мышцах, печени и почках. Белок метаболизируется почками, его время полураспада составляет 2 часа. Астроглиальные клетки – это наиболее многочисленные клетки в мозговой ткани. Они образуют трехмерную сеть, которая является опорным каркасом для нейронов. Увеличение концентрации S‑100 (αβ) и S‑100 (ββ) в СМЖ и плазме является маркером повреждения головного мозга. При раннем определении содержания S‑100 у пациентов с повреждениями мозга концентрация белка отражает степень повреждения мозга. Исследования S‑100 полезны как для мониторинга, так и для определения прогноза течения заболевания.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики