Фотосинтез как основа энергетики биосферы
6.3 САМ-тип метаболизма. Потоки метаболитов в хлоропласт и из него
Суккуленты (роды Crassula, Bryophyllum и др.) также приспособились осуществлять фотосинтез в условиях резко засушливого климата. Для них характерен суточный цикл метаболизма С4-кислот с образованием яблочной кислоты ночью. В соответствии с английским выражением Crassulacean Acid Metabolism (САМ) этот тип фотосинтеза ч
асто сокращенно называют САМ-метаболизм. Устьица этих растений днем обычно закрыты, что предотвращает потерю воды, и открываются ночью. С02 поступает в листья, где при участии содержащейся в цитоплазме ФЕП-карбоксилазы взаимодействует с фосфоенолпируватом, образуя оксалоацетат. Источник ФЕП-карбоксилазы взаимодействует с фосфоенолпируватом, образуя оксалоацетат. Источником ФЕП служит крахмал. То же самое происходит и с С02, который освобождается в клетках в процессе дыхания. Образовавшийся оксалоацетат восстанавливается под действием NADH-зависимой малатдегидрогеназы до яблочной кислоты, которая накапливается в вакуолях клеток листа.
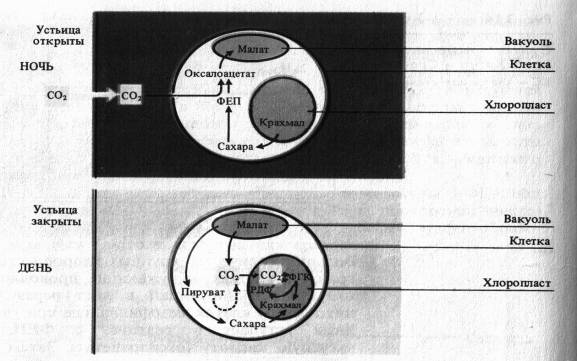
Это приводит к закислению клеточного сока в ночное время. Как и у С4-растений, оксалоацетат может быть источником аспартата, однако этот путь здесь менее выражен.
Днем в условиях высокой температуры, когда устьица закрыты, малат транспортируется из вакуолей в цитоплазму и там декарбоксилируется при участии малатдегидрогеназы декарбоксилирующей (малик-энзима) с образованием С02 и пирувата. С02 поступает в хлоропласты и включается в них в цикл Кальвина, участвуя в синтезе сахаров.
Таким образом, у растений с фотосинтезом по типу толстянковых много общего с С4-путем фотосинтеза. Однако при САМ-метаболизме фиксация С02 с образованием малата (ночью) и декарбоксилирование малата с высвобождением С02 и пирувата (днем) разделены во времени. У С4-растений эти же реакции разграничены в пространстве: первая протекает в хлоропластах мезофилла, вторая — в клетках обкладки. При достаточном количестве воды ряд растений с метаболизмом по типу толстянковых могут вести себя как С3-растения. В свою очередь некоторые растения с С3-путем фотосинтеза при недостатке воды проявляют черты САМ-метаболизма.
7 Фотодыхание, его значение. Сравнение фотодыхания у растений с различными типами метаболизма углерода
В растительных клетках, содержащих хлоропласты, помимо С3- и С4-путей фотосинтеза, осуществляется также фотодыхание, т. е. активируемый светом процесс высвобождения С02 и поглощения 02, который значительно, отличается от «темнового» дыхания митохондрий. Так как при этом первичным продуктом является гликолевая кислота, то этот путь получил название гликолатного. У некоторых С3-растений с малой эффективностью фотосинтеза интенсивность фотодыхания может достигать 50% от интенсивности фотосинтеза.
Фотодыхание у С3-растений обычно усиливается при низком содержании С02 и высоких концентрациях 02. В этих условиях РДФ-карбоксилаза в хлоропластах может функционировать как оксигеназа, катализируя окислительное расщепление рибулозо-1,5-дифосфата на 3-ФГК и 2-фосфогликолевую кислоту, которая затем дефосфорилируется в гликолевую кислоту (рис. 7.1). Молекулы С02 и 02 конкурируют между собой в каталитическом центре РДФ-карбоксилазы: при относительно высоких концентрациях С02 и низких 02 преобладает карбоксилирование, тогда как высокие концентрации 02 и низкое содержание С02 благоприятствуют окислению, а следовательно, и образованию фосфогликолевой кислоты. В том же направлении действует и повышение температуры.
Фотодыхание осуществляется в результате взаимодействия трех органелл — хлоропластов, пероксисом и митохондрий (рис. 7.1). Гликолат из хлоропласта поступает в пероксисому и там окисляется гликолатоксидазой до глиоксилата. Возникающая перекись водорода устраняется каталазой пероксисомы. Глиоксилат аминируется, превращаясь в глицин, причем в качестве донора аминогруппы функционирует глутамат.
Глицин транспортируется в митохондрию. Здесь из двух молекул глицина образуется серин и освобождается С02. Теперь цикл замыкается: серин может снова поступать в пероксисому и там передать свою аминогруппу на пируват. При этом из пирувата возникает аланин, из серина — гидроксипируват, который сразу после этого восстанавливается в глицерат. Затем глицерат может снова попасть в хлоропласты и благодаря фосфорилированию включиться в цикл Кальвина.
Последовательность реакций фотодыхания не обязательно образует цикл. Гликолатный путь С3-растений может завершиться в митохондриях. Конечными продуктами в этом случае является серин и С02. Освобождение С02 объясняет почему нетто-фотосинтез (чистая продуктивность) при интенсивном фотодыхании снижается (рис. 7.1).
У С4-растений С02, выделяющийся в результате фотодыхания, перехватывается в клетках мезофилла, где из ФЕП и С02 образуются оксалоацетат и малат. Затем малат «отдает» свой С02 хлоропластам обкладки, где функционирует цикл Кальвина. В связи с этим становится понятным высокий нетто-фотосинтез С4-растений.
Однако для чего же тогда нужно фотодыхание? Необходимо вспомнить, что гликолатный путь приводит к синтезу глицина и серина, в пероксисомах происходит восстановление NADP+; имеются косвенные данные о том, что в процессе образования серина может генерироваться и АТР. Показано, что С3-растения, помещенные в атмосферу с низким парциальным давлением 02 и высокой концентрацией С02, ведут себя подобно С4-растениям, т. е. имеют низкий уровень фотодыхания.
Из всего сказанного можно сделать вывод, что термин фотодыхание имеет лишь формальный смысл: 02 потребляется, С02 выделяется, однако в функциональном плане к дыханию этот процесс прямого отношения не имеет.
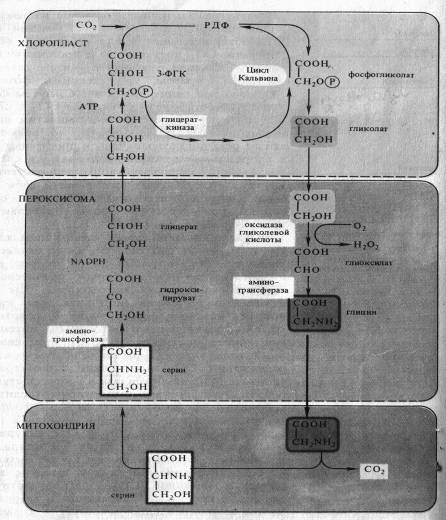
Рис. 7.1
8 Эндогенные механизмы регуляции процесса фотосинтеза
Регуляция фотосинтеза осуществляется на уровне фотохимически активных мембран, хлоропластов, клеток, тканей, органов и целого организма. Все системы регуляции принимают участие в этом процессе.
Процессы в хлоропластах при переходе к фотосинтезу. При освещении листьев хлоропласты уже через несколько минут начинают уменьшаться в объеме, становясь более плоскими (дисковидными). Тилакоиды и граны сдвигаются и уплотняются. Фотоиндуцируемое сокращение хлоропластов объясняется возникновением трансмембранных протонных градиентов и изменением электрического потенциала мембран хлоропластов. Определенная степень сжатия хлоропласта необходима для эффективной работы электронтранспортной цепи и для ее сопряжения с образованием АТР. Транспорт протонов внутрь тилакоидов приводит к подкислению их внутренней полости до рН 5,0—5,5 и одновременному подщелачиванию стромы хлоропласта от рН 7,0 в темноте и до рН 8,0 на свету. Вход ионов Н+ в тилакоиды сопровождается выходом из них в строму ионов Mg2 + .
Появление NADPH, ATP, 02, Mg2+ и изменение рН оказывают прямое и опосредованное влияние на реакции фотосинтетического усвоения С02 в строме. Экспериментально показано, что фиксация С02 изолированными хлоропластами практически не обнаруживается при рН ниже 7,2, но достигает максимальных значений при рН 8,0. Причина этого заключается в изменении активности ряда ферментов, имеющих оптимум рН в слабощелочной среде: рибулозофосфаткиназы (рН 7,9), дегидрогеназы фосфоглицеринового альдегида (рН 7,8), рибулозо-1,5-дифосфаткарбоксилазы (рН от 7,5 до 8,0), фруктозодифосфатазы (рН 7,5 — 8,5). Повышение рН стромы при освещении хлоропласта приводит к значительному связыванию С02 в виде Н2С03 и накоплению НСОз, из которого С02 освобождается с помощью карбоангидразы перед карбоксилированием рибулозо-1,5-дифосфата.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики