Нервная ткань
Постсинаптические потенциалы, возникающие на дендритах, так же невелики, как и рецепторные потенциалы и так же уменьшаются при распространении от синапса до аксонного холмика, где может возникнуть потенциал действия. Кроме того, на пути распространения постсинаптических потенциалов по телу клетки могут оказаться тормозные гиперполяризующие синапсы и потому возможность деполяризации мембраны акс
онного холмика на 10мВ кажется маловероятной. Тем не менее, этот результат регулярно достигается в результате суммации множества небольших постсинаптических потенциалов, возникающих одновременно в многочисленных синапсах, образованных дендритами нейрона с окончаниями аксонов пресинаптических клеток.
Таким образом, объединенный сигнал возникает, как правило, вследствие суммации одновременно образовавшихся многочисленных местных потенциалов. Такая суммация происходит в том месте, где особенно много потенциалзависимых каналов и поэтому легче достигается критический уровень деполяризации. В случае интеграции постсинаптических потенциалов таким местом является аксонный холмик, а суммация рецепторных потенциалов происходит в ближайшем от чувствительных окончаний перехвате Ранвье. Область возникновения объединенного сигнала называется интегративной или триггерной (от англ. trigger - спусковой крючок).
Проведение потенциала действия
Как уже говорилось, амплитуда входных сигналов пропорциональна силе подействовавшего стимула или количеству выделившегося в синапс нейромедиатора - такие сигналы называют градуальными. Их длительность определяется длительностью стимула или присутствия медиатора в синаптической щели. Амплитуда и длительность потенциала действия от этих факторов не зависят: оба этих параметра всецело определяются свойствами самих клеток. Следовательно, любая комбинация входных сигналов, любой вариант суммации, при единственном условии деполяризации мембраны до критического значения, вызывает один и тот же стандартный образец потенциала действия в триггерной зоне. Он всегда имеет максимальную для данной клетки амплитуду и примерно одинаковую длительность, сколько бы раз ни повторялись вызывающие его условия.
Возникнув в интегративной зоне, потенциал действия быстро распространяется по мембране аксона. Это происходит благодаря появлению локального электрического тока. Поскольку деполяризованный участок мембраны оказывается иначе заряженным, чем соседствующий с ним, между полярно заряженными участками мембраны возникает электрический ток. Под действием этого локального тока деполяризуется до критического уровня соседний участок, что вызывает появление потенциала действия в нем. В случае миелинизированного аксона таким соседним участком мембраны является ближайший к триггерной зоне перехват Ранвье, затем следующий, и потенциал действия начинает «перепрыгивать» от одного перехвата к другому.
Разные нейроны могут многим отличаться друг от друга, но возникающие в них потенциалы действия различить очень трудно, а в большинстве случаев и невозможно. Это в высшей степени стереотипный сигнал у самых разных клеток: сенсорных, интернейронов, моторных. И это свидетельствует о том, что сам потенциал действия не содержит никаких сведений о природе породившего его стимула. О силе стимула свидетельствует частота возникающих потенциалов действия, а определением природы стимула занимаются специфические рецепторы и хорошо упорядоченные межнейронные связи.
Таким образом, возникший в триггерной зоне потенциал действия быстро распространяется по ходу аксона к его окончанию. Это передвижение связано с образованием локальных электрических токов, под влиянием которых потенциал действия как бы заново возникает в соседнем участке аксона. Параметры потенциала действия при проведении по аксону нисколько не меняются, что позволяет передавать информацию без искажений. Если аксоны нескольких нейронов оказываются в общем пучке волокон, то по каждому из них возбуждение распространяется изолированно.
Выходной сигнал
Выходной сигнал адресуется другой клетке или одновременно нескольким клеткам и в подавляющем большинстве случаев представляет собой выделение химического посредника - нейротрансмиттера или медиатора В пресинаптических окончаниях аксона заранее запасенный медиатор хранится в синаптических пузырьках, которые накапливаются в специальных участках - активных зонах. Когда потенциал действия добирается до пресинаптического окончания, содержимое синаптических пузырьков путем экзоцитоза опорожняется в синаптическую щель.
Химическими посредниками передачи информации могут служить разные вещества: небольшие молекулы, как, например, ацетилхолин или глутамат, либо достаточно крупные молекулы пептидов - все они специально синтезируются в нейроне для передачи сигнала. Попов в синаптическую щель, медиатор диффундирует к постсинаптической мембране и присоединяется к ее рецепторам. В результате связи рецепторов с медиатором изменяется ионный ток через каналы постсинаптической мембраны, а это приводит к изменению значения потенциала покоя постсинаптической клетки, т. е. в ней возникает входной сигнал - в данном случае постсинаптический потенциал.
Таким образом, почти в каждом нейроне, независимо от его величины, формы и занимаемой в цепи нейронов позиции, можно обнаружить 4 функциональные области: локальную рецептивную зону, интегративную, зону проведения сигнала и выходную или секреторную зону.
Глия
Во всех органах человеческого тела, кроме мозга, функционирующие клетки удерживаются вместе межклеточным веществом соединительной ткани В нервной системе эту роль выполняет глия (от греч. глия - клей), клетки которой образуются из общих с нейронами предшественниц на раннем этапе развития мозга. Глия создает опору для нейронов, объединяет отдельные элементы нервной системы, но, в то же время, изолирует друг от друга разные группы нейронов, а также большую часть их аксонов. Тем самым она формирует структуру мозга. Численность клеток глии превышает количество нейронов в мозгу приблизительно в 10 раз. Эти клетки отличаются друг от друга по внешнему виду и по выполняемой функции.
Клетки ейроглии
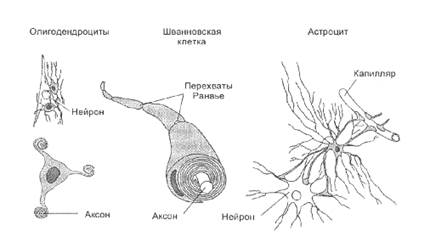
Самыми распространенными среди клеток глии являются астроциты, например, в мозолистом теле они составляют 1/4 всех клеток глии. У астроцита неправильной, звездчатой формы тело с многочисленными и относительно длинными отростками, одни из которых направлены к нейронам, а другие - к кровеносным капиллярам. Эти отростки расширяются на концах, образуя т. н. а строцитарную ножку. На поверхности капилляра отростки соседних астроцитов плотно смыкаются друг с другом и практически полностью обвертывают кровеносный сосуд. Подобная изоляция сосуда является одним из способов формирования гематоэнцефалического барьера - граници между кровью и нервной тканью, закрытой для многих находящихся в крови веществ.
Другие отростки астроцита почти целиком обвертывают тела нейронов. Если нейрон возбуждается длительно, вокруг него повышается концентрация ионов калия, а это может уменьшить возбудимость соседних нейронов. Астроциты предупреждают такую возможность, поглощая излишки калия, - тем самым они выполняют функцию буфера. Некоторые клетки глии при этом деполяризуются, а поскольку они связанны между собою щелевыми контактами, между деполяризованными и находящимися в покое клетками возникает ток. Это, однако, не приводит к возбуждению, так как в мембране клеток глии очень мало потенциалзависимых каналов для натрия или кальция. Несмотря на то, что повышение концентрации ионов калия у астроцитов изменяет некоторые их свойства, в настоящее время нет достаточных оснований считать их прямыми участниками переноса нервных импульсов.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики