ДНК. Основы генетического материал
Химически ДНК представляют собой высокомолекулярные полимеры монодезоксирибонуклеотидов (мононуклеотидов), являющиеся мономерами, из которых построены молекулы
ДНК. Каждый мононуклеотид ДНК состоит из остатков фосфорной кислоты, 2-П-дезоксирибозы и пуринового или пиримидинового азотистого основания. Углеводно-фосфатный остаток одинаков во всех мономерах ДНК, азотистое основание же может быт
ь представлено аденином (А), гуанином (Г), цитозином (Ц) или тимином (Т). В ДНК разных организмов имеется некоторое количество так называемых, минорных оснований, например 5-метил-цитозина, частично заменяющего цитозин. У высших животных и человека содержание этого основания достигает 1,5%, у высших растений 5—7% , у бактерий — не более 0,6% . В ДНК бактерий встречается также 6-метиладенин и иногда другие метилированные азотистые основания. В ДНК Т-четных бактериофагов (Т2, Т4 и Т6) цитозин полностью замещен 5-оксиметилцитозином, в ДНК вирусов SP01 и SP8 тимин замещен 5-оксиметилурацилом, а у фага PBS1 — урацилом.
В мононуклоотидах 2-П-дезокси-рибоза присоединена гликозидной связью через первый углеродный атом к атому азота в 9-м положении пуринового основания (аденина или гуанина) или в 3-м положении пиримидинового основания (цитозина или тимина). Остаток фосфорной кислоты присоединен эфирной связью к 5'-му или З'-му атому углерода дезоксирибозы. Таким образом, мононуклеотидные остатки соединены между собой через фосфорную кислоту, которая соединена с 5'-С-атомом дезоксирибозы одного нуклеотида и с 3'-С-атомом дезоксирибозы соседнего нуклеотида и т. д. (схема 1).
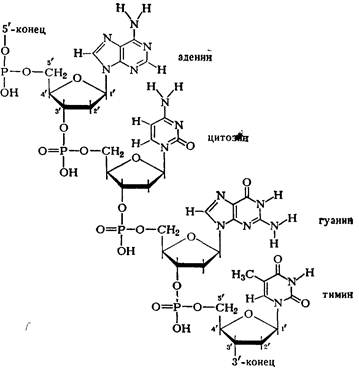
Схема 1. Соединение нуклеотидов в молекуле ДНК.
ДНК из различных источников отличаются друг от друга по соотношению входящих в их состав азотистых оснований, то есть по нуклеотидному составу, однако нуклеотидный состав всех ДНК подчиняется определенным закономерностям — правилам Чаргаффа, согласно которым:
1) число молекул аденина равно числу молекул тимина; 2) число молекул гуанина равно числу молекул цитозина; 3) число молекул пуриновых оснований равно числу молекул пиримидиновых оснований;
4) число 6-аминогрупп в молекуле ДНК равно числу 6-кетогрупп, то есть сумма аденин + цитознн равна сумме гуанин + тимин. Записав правила Чаргаффа буквенными обозначениями, получим следующие выражения: 1) А - Т; 2) Г - Ц; 3) А + Г = Т + Ц; 4) А + Ц = Г + Т. Эти правила сохраняют силу и в том случае, если приведенные азотистые основания замещены их метилированными или другими производными (минорными основаниями). Таким образом, нуклеотидный состав ДНК характеризуется молярным отношением ![]() (фактором специфичности) или процентом ГЦ-пар, т.е.
(фактором специфичности) или процентом ГЦ-пар, т.е. ![]() . Величинаэтого показателя одинакова для ДНК различных органов и тканей одного организма и практически не отличается у разных видов животных и растений в пределах одного класса. Она достаточно близка у высших растений и животных (позвоночных) — от 0,55 до 0,93. У бактерий, по данным А. С. Спирина и А. Н. Белозерского, величина фактора специфичности колеблется от 0,35 до 2,73 или от 26,8 до 74,2% ГЦ-пар.
. Величинаэтого показателя одинакова для ДНК различных органов и тканей одного организма и практически не отличается у разных видов животных и растений в пределах одного класса. Она достаточно близка у высших растений и животных (позвоночных) — от 0,55 до 0,93. У бактерий, по данным А. С. Спирина и А. Н. Белозерского, величина фактора специфичности колеблется от 0,35 до 2,73 или от 26,8 до 74,2% ГЦ-пар.
Рентгеноструктурный анализ ДНК показал, что пуриновые и пиримидиновые основания нуклеотидных остатков ДНК лежат в одной плоскости, перпендикулярной продольной оси молекулы, тогда как циклы дезоксирибозы находятся в плоскости, почти перпендикулярной той, в которой лежат циклы оснований. Расстояния между азотистыми основаниями отдельных нуклеотидов составляют 3,4 А. В соответствии с этими данными и с правилами Чаргаффа Дж. Уотсон и Ф. Крик построили модель молекулы ДНК (схема 2). Дальнейшие исследования подтвердили их правоту. Установление строения молекулы ДНК явилось крупнейшим открытием в области молекулярной биологии. Согласно модели Уотсона - Крика, молекула ДНК представляет собой двойную спираль, построенную из двух полинуклеотидных цепочек, направленных антипараллельно, то есть если в одной цепочке остаток фосфорной кислоты связывает отдельные нуклеотиды от 5'- к 3'-С-атомам снизу вверх, то в другой цепочке эти связи направлены сверху вниз. Каждая цепочка состоит из углеводно-фосфорного скелета, присоединенные к углеводному компоненту азотистые основания ориентированы внутрь и соединены между собой попарно водородными связями, а именно А– с Т и Г – с Ц. Аденин с тимином соединены двумя Н-связями, тогда как гуанин с цитозином соединены еще третьей водородной связью (схема 3). Двойная спираль закручена вправо, причем полному витку спирали соответствуют 10 пар нуклеотидных остатков, занимающих расстояние в 34 А,— В-форма. В-форма устойчива в среде с высокой влажностью (97% насыщенного пара). Вся молекула ДНК представляет собой жесткий, неветвящийся линейный полимер. В условиях низкой влажности (с 76% насыщения) двойная спираль ДНК принимает А-форму, в которой полный виток спирали занимает расстояние в 28 А, причем меняется также положение плоскости, в которой расположены азотистые основания, и число оснований на полный виток (один виток содержит 11 нуклеотидов).
В хроматине ДНК образует комплексы с гистонами. Такие нуклеогистоны находятся в сверхспирализованном состоянии, причем суперспираль имеет радиус 50 А и расстояние между витками 120 А. В хромосомах и частично в хроматине такие суперспирали дезоксирибонуклеопротеида закручены в спирали высших порядков с диам. 250 и 500 А.
Молекулярный вес (масса) ДНК неодинаков и зависит от источников получения образца ДНК. Помимо этого, даже при самых тщательных и щадящих процедурах выделения ДНК подвергается некоторой деградации и ее молекулярный вес может быть ниже, чем в клетках. Препараты, получаемые современными методами из тканей животных н растений, имеют мол. вес 6-106—10-106, однако истинный мол. вес ДНК животных и растений, как показывают методы определения мол. веса по вязкости и по длине молекул (lA двуспиральной ДНК в В-форме соответствует 197 единицам молекулярного веса), значительно выше и может достигать десятков миллиардов. Таким образом. молекулы ДНК хромосом являются самыми крупными молекулами из всех известных биополимеров.
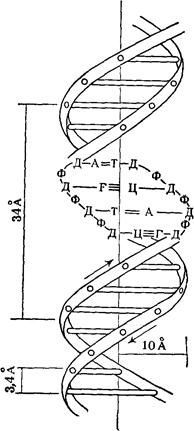
Схема 2. Двойная спираль молекулы ДНК (модель Уотсона—Крика): А — аденин; Т — тимин, Г — гуанин; Ц — цитозин; Д — дсзоксирибоза; Ф — фосфат; 34 А — величина витка, спирали; 10 А — радиус спирали; 3, 4 А — расстояние между нуклеотидами; стрелки указывают направление витка спирали.
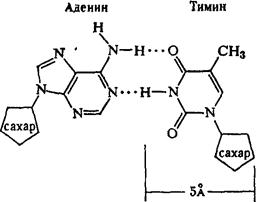
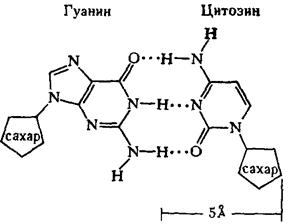
Схема 3. Соединение пуриновых и пиримидиновых оснований в молекуле ДНК (точками обозначены водородные связи).
У некоторых вирусов, например у бактериофагов Ф Х174, fd и М13, ДНК представлена одной полинуклеотидной цепью, замкнутой в кольцо и имеющей сравнительно небольшой мол. вес — 1,7-106. У большинства вирусов ДНК представляет собой двойную спираль, линейную пли замкнутую в кольцо; нередко такие формы переходят друг в друга, причем эти молекулы имеют так называемые «липкие концы», содержащие однонитчатые комплементарные друг другу нуклеотидные последовательности, при помощи которых молекула замыкается в кольцо. Для ДНК характерно сильное поглощение в ультрафиолетовой части спектра при длине волны около 260 нм. Удельное поглощение высокополимерной ДНК в растворе, содержащем выше 10-3 М NaCI, при рН 7,0 составляет около 6000 на 1 г-атом фосфора. ДНК сравнительно легко деполимеризуются под действием некоторых химических соединений, ультразвука, ионизирующей и ультрафиолетовой радиации; нагревание ДНК с разведенными минеральными кислотам приводит к отщеплению пуринов (аденина и гуанина) и образованию «апуриновой кислоты», содержащей только пиримидиновые основания. Нагревание растворов ДНК, а также их подщелачивание и т.п. вызывают денатурацию ДНК, заключающуюся в плавлении двойной спирали (разрыве водородных и гидрофобных связей) и расхождении полинуклеотидных цепочек. Денатурация сопровождается понижением вязкости раствора и повышением поглощения в ультрафиолете, по чему можно контролировать этот процесс. Температура плавления (температура, при которой денатурирована половина ДНК) тем выше, чем больший процент ГЦ-пар содержится в ДНК; этот показатель может служить для определения нуклеотидного состава ДНК. Установлено, что (не линейно связана с составом ДНК: 1° соответствует 2,5 молярным % ГЦ-пар. Гомогенные препараты ДНК (например, вирусной ДНК) характеризуются плавлением с резким переходом, тогда как гетерогенные препараты дают сравнительно широкую зону плавления, что может служить мерой гетерогенности ДНК. При быстром охлаждении после денатурации ДНК не восстанавливает своих нативных свойств, однако при медленном охлаждении полинуклеотидные цепочки реассоциируются по принципу комплементарности и таким образом происходит ренатурация молекул ДНК. При медленном охлаждении денатурированной ДНК в присутствии РНК полинуклеотидные нити ДНК и РНК могут ассоциироваться но принципу комплементарности пар гуанина с цитозином и аденина с урацилом (вместо тимина), образуя двунитчатые гибриды ДНК — РНК. Метод гибридизации широко применяется для исследования комплементарности и структуры двух типов нуклеиновых кислот, а также ДНК из разных источников. Изучение ренатурации ДНК показало, что ДНК высших организмов содержат повторяющиеся последовательности, которые можно разделить на очень часто повторяющиеся последовательности и относительно часто повторяющиеся. Кроме того, имеются и уникальные последовательности. К повторяющимся последовательностям, по-видимому, относятся регуляторные гены, а также гены, кодирующие рибосомные РНК, транспортные РНК и гистоны. Структурные гены, как правило, относятся к уникальным последовательностям, что доказано для таких активных генов, как гены глобина в эритробластах и гены фиброина в шелкоотделительной железе шелкопряда. У низших организмов (прокариотов) — вирусов и бактерий, а также в митохондриях ДНК не содержит или почти не содержит повторяющихся последовательностей. В ДНК ряда организмов обнаружены участки, в каждой из нуклеотидных цепей которых имеются последовательности оснований, повторяющиеся далее, но в обратном порядке. Поскольку такие последовательности читаются одинаково с обоих концов, как, например, слово «потоп», они получили название палиндромов. Палиндромы в ДНК и в синтезированных на их матрице РНК могут образовывать крестообразные структуры, физиологическую роль которых, возможно, связана с инициацией (началом) синтеза РНК или белков.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики