Полимеразная цепная реакция
В конечном счете, если эти осложнения никак более не удается преодолеть, можно при секвенировании использовать внутренние праймеры, отжигающиеся только на интересующей последовательности.
Еще одно неудобство, вызванное прямым секвенированием продуктов ПЦР, обусловлено способностью двух цепей амплифицированного фрагмента к быстрой реассоциации, что мешает отжигу секвенационного праймера на к
омплементарной последовательности или блокирует реакцию достройки комплекса затравка — матрица. Устранить это препятствие можно, применяя один из вариантов стандартного метода секвенирования двухцепочечной ДНК или модифицируя ПЦР с целью получения одноцепочечных продуктов.
3.1 Секвестрование двухцепочечных образцов
Для приготовления двухцепочечных образцов, используемых при секвенировании, выберите одну из двух методик.
1. Денатурируйте образец NaOH, перенесите в лед, нейтрализуйте реакцию, после чего быстро осадите ДНК- Растворите ДНК в буфере, содержащем секвенационный праймер, при необходимой температуре отжига.
2. Денатурируйте образец нагреванием, затем быстро перенесите в спиртовую баню, охлажденную до температуры сухого льда, замедлив таким образом процесс реассоциации цепей. Добавьте секвенационный праймер и нагрейте до необходимой температуры отжига.
Для секвенирования двухцепочечных плазмидных ДНК подходят оба способа. Секвенировать продукты ПЦР по этим инструкциям гораздо труднее, так как короткие линейные фрагменты ДНК более склонны к реассоциации, чем двухцепочечные кольцевые плазмиды.
3.2 Секвенирование одноцепочечных фрагментов ДНК
Затруднения, связанные с реассоциацией цепей фрагмента, можно преодолеть, если приготовить одноцепочечные образцы ДНК путем разделения цепей в геле или синтезировать их в самой реакции ПЦР. Для разделения цепей фрагментов длиной не менее 500 п. н. могут быть с успехом использованы соответствующие агарозные гели. В случае более коротких фрагментов такой подход не эффективен. Однако недавно мы разработали метод, позволяющий синтезировать одноцепочечные ДНК в ходе реакции ПЦР.
Эта процедура предусматривает использование неэквивалентных количеств двух праймеров для амплификации, что позволяет синтезировать за первые 20-25 циклов двухцепочечную ДНК и за последующие 5-10 циклов в условиях недостатка одного из праймеров — одноцепочечную ДНК.
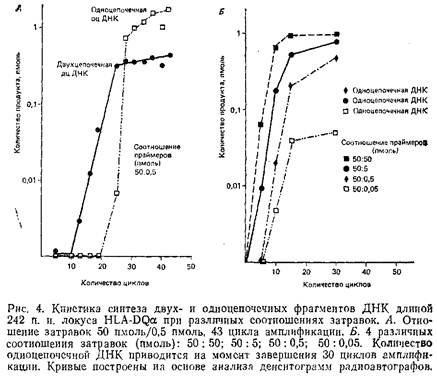
Рис. 4, А демонстрирует накопление дцДНК и оцДНК в ходе обычной амплификации геномной последовательности, использующей исходное соотношение затравок 50:0,5 пмоль в 100 мкл реакционной смеси. Как и ожидалось, количество дцДНК возрастает экспоненциально до момента практического исчезновения из реакционной смеси одного из праймеров, после чего количество этой ДНК нарастает очень медленно. Фракция оцДНК появляется, начиная с 25-го цикла, с момента, когда запас лимитирующей затравки почти полностью исчерпан. После короткой начальной стадии быстрого роста оцДНК накапливается линейно, чего следует ожидать в присутствии одного праймера.
Разные соотношения праймеров дадут в принципе сходную картину образования оцДНК. Различные неравные отношения 50:5, 50:0,5, 50:0,05 дают за 30 циклов полимеразной реакции количество оцДНК, превышающее количество синтезированной дцДНК. Как правило, соотношение 50:0,5 позволяет получить после 30 циклов ПЦР 1-5 пмоль оцДНК. Образованная оцДНК может быть затем секвенирована с использованием лимитирующего праймера ПЦР или внутреннего праймера по обычной схеме химической или энзиматической методики секвенирования. Синтезированные фрагменты будут иметь гомогенный 5'-конец и в различной степени усеченный 3'-конец вследствие преждевременной терминами синтеза. Однако при использовании некоторых секвенационных затравок только полная оцДНК. может служить образцом для секвенирования.

1. Проведите реакцию ПЦР по уже описанной методике, изменив только количества используемых праймеров на 50 пмоль и 0,5 пмоль в 100 мкл реакционной смеси. Проведите 30— 35 циклов реакции. Если необходимо секвенировать обе цепи, приготовьте также образец с обратным соотношением объема используемых затравок.
2. После завершения ПЦР смешайте 100 мкл реакционной смеси с 2 мл дистиллированной воды, заполните этой смесью микроконцентратор Centricon 30 и для удаления избытка dNTP и компонентов буфера отцентрнфугируйте при 5000 об/мин в роторе с фиксированным наклоном пробирок.
3. Высушите 10 из 40 мкл концентрата и растворите в 10 мкл секвенационного буфера, содержащего 1 пмоль секвенационного праймера.
4. Прогрейте смесь праймер — матрица при 65 °С в течение 2 мин, затем оставьте охлаждаться до 30 °С в течение 20 мин.
5. Добавьте 1 мкл 100 мМ DDT, 2 мкл смеси для включения метки, разведенной в соотношении 1/100, 0,5 мкл dATP, 10 мкКи/мкл, 2 мкл ДНК-полимеразы Т7.
6. Внесите 3,5 мкл приготовленной смеси в каждую из 4 пробирок, содержащих 2,5 мкл смеси дидезоксирибонуклеозидов, и инкубируйте 5 мин при 37°С.
Остановите реакцию добавлением 4 мкл 95%-ного формамида, 20 мм ЭДТА; прогрейте в течение 2 мин при 75°С и нанесите на секвенационный гель.
Заключение
ПЦР открывает возможность быстрого синтеза миллионов копий индивидуальной последовательности ДНК, что значительно упрощает последующий ее анализ.
Поскольку в результате цепной амплификации образуются идентичные фрагменты специфичной ДНК, сразу же может быть осуществлено клонирование или секвенирование продуктов реакции. Матрицей для ПЦР-амплификации могут служить как геномная ДНК, так и кДНК, синтезированная путем обратной транскрипции РНК- Чувствительность метода позволяет обнаружить и амплифицировать единственную молекулу ДНК- При автоматизации ПЦР и наличии нерадиоактивных материалов для мечения ДНК полнмеразная цепная реакция обещает стать в будущем стандартной молекулярно-биологической процедурой. Можно предположить, что велика будет и ее роль в диагностике наследственных и инфекционных заболеваний.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики