Структурно-функциональная организация генетического материала
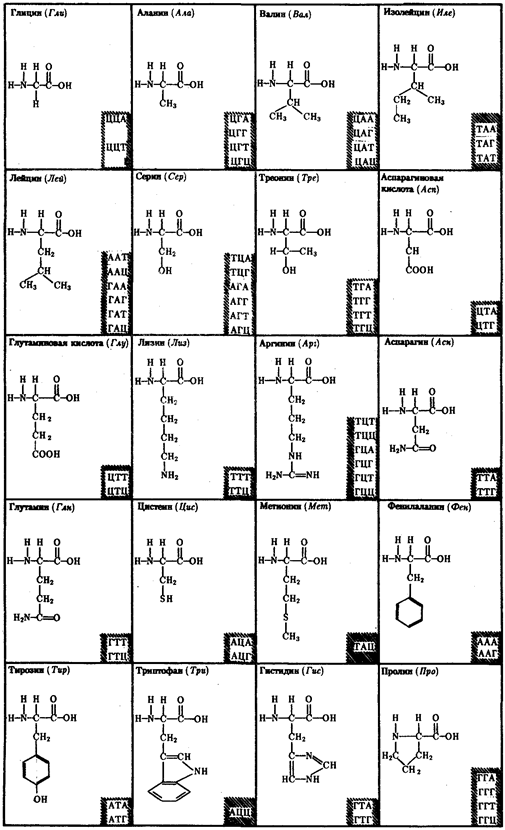
Рис.6. Аминокислоты и кодирующие их триплеты ДНК
Другие особенности являются специфичными для различных видов организмов. У дрожжей триплет ГАТ и, возможно, все семейство ГА кодирует вместо аминокислоты лейцина треонин. У млекопитающих триплет ТАГ имеет то же значение, что и ТАЦ, и кодирует аминокислоту метиони
н вместо изолейцина. Триплеты ТЦГ и ТЦЦ в ДНК митохондрий некоторых видов не кодируют аминокислот, являясь нонсенс-триплетами. Наряду с триплетностью, вырожденностью, специфичностью и универсальностью важнейшими характеристиками генетического кода являются его непрерывность и неперекрываемость кодонов при считывании. Это означает, что последовательность нуклеотидов считывается триплет за триплетом без пропусков, при этом соседние триплеты не перекрывают друг друга, т.е. каждый отдельный нуклеотид входит в состав только одного триплета при заданной рамке считывания (рис.3.7). Доказательством неперекрываемости генетического кода является замена только одной аминокислоты в пептиде при замене одного нуклеотида в ДНК. В случае включения нуклеотида в несколько перекрывающихся триплетов его замена влекла бы за собой замену 2-3 аминокислот в пептидной цепи.
Рис.7. Непрерывность и непререкаемость генетического кода при считывании наследственной информации.
Цифрами обозначены нуклеотиды
4.2 Свойства ДНК как вещества наследственности и изменчивости
4.2.1 Самовоспроизведение наследственного материала. Репликация ДНК
Одним из основных свойств материала наследственности является его способность к самокопированию - репликация. Это свойство обеспечивается особенностями химической организации молекулы ДНК, состоящей из двух комплементарных цепей. В процессе репликации на каждой полинуклеотидной цепи материнской молекулы ДНК синтезируется комплементарная ей цепь. В итоге из одной двойной спирали ДНК образуются две идентичные двойные спирали. Такой способ удвоения молекул, при котором каждая дочерняя молекула содержит одну материнскую и одну вновь синтезированную цепь, называют полуконсервативным.
Для осуществления репликации цепи материнской ДНК должны быть отделены друг от друга, чтобы стать матрицами, на которых будут синтезироваться комплементарные цепи дочерних молекул.
Инициация репликации осуществляется в особых участках ДНК, обозначаемых ori (от англ. origin - начало). Они включают последовательность, состоящую из 300 нуклеотидных пар, узнаваемую специфическими белками. Двойная спираль ДНК в этих локусах разделяется на две цепи, при этом, как правило, по обе стороны от точки начала репликации образуются области расхождения полинуклеотидных цепей - репликационные вилки, которые движутся в противоположных от локуса ori направлениях. Между репликационными вилками образуется структура, называемая репликационным глазком, где на двух цепях материнской ДНК образуются новые полинуклеотидные цепи (рис 8, А).
С помощью фермента геликазы, разрывающего водородные связи, двойная спираль ДНК расплетается в точках начала репликации. Образующиеся при этом одинарные цепи ДНК связываются специальными дестабилизирующими белками, которые растягивают остовы цепей, делая их азотистые основания доступными для связывания с комплементарными нуклеотидами, находящимися в нуклеоплазме. На каждой из цепей, образующихся в области репликационной вилки, при участии фермента ДНК-полимеразы осуществляется синтез комплементарных цепей (рис 8, Б).
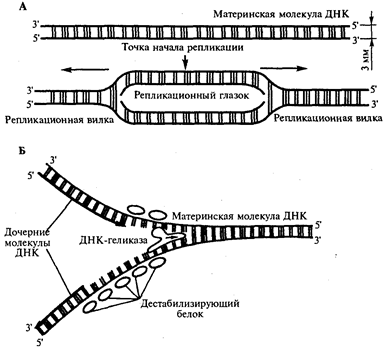
Рис.8. Область начала репликации. Репликационная вилка
А. Образование репликационного глазка.
В. Область репликационной вилки в молекуле ДНК
В процессе синтеза репликационные вилки движутся вдоль материнской спирали в противоположных направлениях, захватывая все новые зоны.
Разделение спирально закрученных цепей родительской ДНК ферментом геликазой вызывает появление супервитков перед репликационной вилкой. Это объясняется тем, что при расхождении каждых 10 пар нуклеотидов, образующих один виток спирали, родительская ДНК должна совершить один полный оборот вокруг своей оси. Следовательно, для продвижения репликационной вилки вся молекула ДНК перед ней должна была бы быстро вращаться, что потребовало бы большой затраты энергии. В действительности это не наблюдается благодаря особому классу белков, называемых ДНК-топоизомеразами. Топоизомераза разрывает одну из цепей ДНК, что дает ей возможность вращаться вокруг второй цепи. Это ослабляет накопившееся напряжение в двойной спирали ДНК (рис.9).
К высвобождающимся водородным связям нуклеотидных последовательностей разделенных родительских цепей присоединяются свободные нуклеотиды из нуклеоплазмы, где они присутствуют в виде дезоксирибонуклеозидгрифосфатов: дАТФ, дГТФ, дЦТФ, дТТФ. Комплементарный нуклеозидтрифосфат образует водородные связи с определенным основанием материнской цепи ДНК. Затем при участии фермента ДНК-полимеразы он связывается фосфодиэфирной связью с предшествующим нуклеотидом вновь синтезируемой цепи, отдавая при этом неорганический пирофосфат (рис.10).
Поскольку ДНК-полимераза присоединяет очередной нуклеотид к ОН-группе в 3'-положении предшествующего нуклеотида, цепь постепенно удлиняется на ее 3'-конце.
Особенностью ДНК-полимеразы является ее неспособность начать синтез новой полинуклеотидной цепи путем простого связывания двух нуклеозидтрифосфатов: необходим 3'-ОН-конец какой-либо полинуклеотидной цепи, спаренной с матричной цепью ДНК, к которой ДНК-полимераза может лишь добавлять новые нуклеотиды. Такую полинук-леотидную цепь называют затравкой или праймером.
Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы (рис.11). Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3'-ОН-конец.
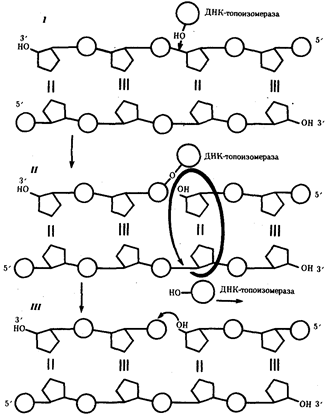
Рис.9. Разрыв одной из цепей ДНК с помощью фермента ДНК-топоизомеразы: I - ДНК-топоизомераза образует ковалентаую связь с одной из фосфатных групп ДНК (верхняя цепь); II - в результате разрыва фосфодиэфирной связи в одной полинуклеотидной цепи вокруг соответствующей ей связи другой цепи осуществляется вращение, которое снимает напряжение, вызванное расхождением двух цепей ДНК в области репликационной вилки; III - после снятия напряжения в спирали ДНК происходит спонтанное отделение ДНК-топоизомеразы и восстановление фосфодиэфирной связи в цепи ДНК
Способность ДНК-полимеразы осуществлять сборку полинуклеотида в направлении от 5' - к 3' - концу при антипараллельном соединении двух цепей ДНК означает, что процесс репликации должен протекать на них по-разному. Действительно, если на одной из матриц (3' → 5') сборка новой цепи происходит непрерывно от 5' - к 3'-концу и она постепенно удлиняется на 3'-конце, то другая цепь, синтезируемая на матрице (5' → 3'), должна была бы расти от 3' - к 5'-концу. Это противоречит направлению действия фермента ДНК-полимеразы.
Другие рефераты на тему «Биология и естествознание»:
Поиск рефератов
Последние рефераты раздела
- Влияние экологических факторов на разнообразие моллюсков разнотипных искусственных и естественных водоемов
- Влияние экологии водоемов на биологическое разнообразие фауны
- Влияние фтора и фторосодержащих соединений на здоровье населения
- Влияние факторов внешней среды на микроорганизмы
- Влияние физической нагрузки на уровень адренокортикотропного гормона, адреналина, кортизола, кортикостерона в сыворотке крови спортсменов
- Временные аспекты морфогенетических процессов. Эволюция путем гетерохронии
- Вопросы биоэтики