Разработка методов биотехнологического получения белков, аминокислот и нуклеозидов, меченных дейтерием и изотопом углерода 13С с высокими степенями изотопного обогащения
Как и в случае с экзогенными аминокислотами, низкие степени включения изотопа углерода 13С в остатки Leu при росте на 1 об.% 13СНзОН обусловлены ауксотрофностью бактерий в этой аминокислоте.
Таким образом, нам удалось достичь максимальных уровней включения стабильных изотопов в суммарные белки биомассы метилотрофных бактерий. Именно поэтому мы посчитали возможным использовать гидролизаты их
биомассы для биосинтеза других изотопно - меченных БАС.
5. ИССЛЕДОВАНИЕ ВОЗМОЖНОСТИ ИСПОЛЬЗОВАНИЯ ГИДРОЛИЗАТОВ БИОМАССЫ МЕТИЛОТРОФНЫХ БАКТЕРИЙ В. methylkumв КАЧЕСТВЕ СУБСТРАТОВ ДЛЯ ПОЛУЧЕНИЯ [1',3',4',2,8-D5]-ИНОЗИНА.
Получение [1',3',4',2,8-D5]- инозина. В следующих экспериментах было апробировано использование дейтеро-компонентов биомассы метилотрофных бактерий, полученных в условиях многоступенчатой адаптации к тяжёлой воде для синтеза высокодейтерированных нуклеозидов (на примере инозина). [1,3',4',2,8-D5]-инозин был получен биосинтетически за счёт использования штамма-продуцента В. subtilis и выделен из культуральной жидкости по методике, включающей адсорбцию инозина на активированном угле, десорбцию спиртово-аммиачным раствором и перекристаллизацию из метанола. ТСХ инозина, с детекцией при 249 нм показала наличие в анализируемом образце единственного пятна с Rf = 0,55, -соответствующего по подвижности чистому инозину.
Особенности разработанного метода получения [1,3',4',2,8-D5]-инозина заключаются в следующих аспектах:
1. В способности высокоактивного штамма В. subtilis к росту и биосинтезу инозина на средах, содержащих максимальные концентрации тяжёлой воды;
2. Замене глюкозы и аминокислот, необходимых для роста этого штамма-ауксотрофа на гидролизаты дейтеро-биомассы В. methylicutn. При последующих ферментациях в качестве источника ростовых факторов можно использовать ту же дсйтеро-биомассу метилотрофных бактерий, либо биомассу самого штамма-продуцента, содержащую в своем составе соединения, которые могут служить источниками углерода и ростовых факторов;
3. В практически полном отсутствии отходов: согласно схеме, дейтеро-биомасса базового штамма, после гидролиза в 6 н. DC1 возвращается в цикл в качестве ростовых факторов;
4. В высокой степени изотопного обогащения дейтерий-мсченного инозина (62,5% атомов водорода в молекуле замещены на дейтерий);
5. В высоких выходах (3,9 г/л) меченного продукта.
Исследование уровня дейтерированности инозина. Места локализации дейтерия в молекуле инозина, были исследованы с помощью масс-спектрометрии FAB и спектроскопии ПМР (см. схему).
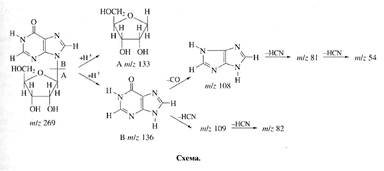
При анализе степени дейтерированности инозина учитывались следующие аспекты. Во-первых, вследствие того, что протоны в C1-C's положениях рибозной части молекулы инозина могли происходить из глюкозы, мы предположили,что характер биосинтетического включения дейтерия в рибозную часть молекулы инозина определяется, в основном, функционированием ряда процессов гексозо-моно-фосфатного (ГМФ) шунта, связанных непосредственно с ассимиляцией глюкозы и других сахаров. Во-вторых, многочисленные обменные процессы и внутримолекулярные перегруппировки, происходящие с участием тяжёлой воды могли также привести к специфическому включению метки по определенным позициям в молекуле инозина. Такими доступными позициями в молекуле инозина признаны, прежде всего, гидроксильные протоны -ОН и протоны при гетероатомах -NH (последние могут обмениваться на дейтерий в тяжёлой воде за счет кето-енольной таутомерии). Три атома дейтерия в рибозном остатке молекулы инозина могли происходить за счет функционирования многочисленных реакций ГМФ-шунта, два атома дейтерия а гипоксантинс также могли синтезироваться de novo (схема).
6. РАЗРАБОТКА СПОСОБОВ БИОСИНТЕТИЧЕСКОГО ПОЛУЧЕНИЯ ДЕЙТЕРИЙ-МЕЧЕНHOI О БАКТЕРИОРОДОПСИНА
Получение дейтерии-меченного бактериородопсина.
В качестве другой модельной системы для введения стабильной изотопной метки в белки, использовали бактериородопсин (bR), синтезируемый в клеточной мембране H. halobium ET 1QOL (рис.6). Для включения дейтериевой метки в bR использовали два принципиально отличных подхода: сайт-специфическое введение отдельных аминокислот: L-[2,3,4,5,6-D5]-Phe, L-[3,5-D2]-Tyr и L-[2,4,5,6,7-D5]-Trp в bR и униформное мечение бактериородопсина дейтерием путем выращивания П. halobium ET 1001 на среде, содержащей 99,9 ат.% тяжёлую воду и дейтеро-гидролизаты В. methylicum.
Бактериородопсин выделяли из пурпурных мембран Н. halobium ET 1001 солюбилизацией в 0,5 %-ном растворе додецилсульфата натрия (ДСН) с последующим осаждением белка метанолом. Гомогенность очищенного bR была потверждена электрофорезом в 12,5%-ном полиакриламидном геле в присутствии 0,1% ДСН.
ОФ ВЭЖХ метиловых эфиров Dns-, и Z-производных аминокислот, полученных после гидролиза bR в 4 н. Ва(ОН)2 или 6 н. DCI (3 масс.% фенола, в тяжёлой воде) показала высокие степени хроматографической чистоты выделенных аминокислот и отсутствие примесей небелковой природы в гидролизатах bR. Согласно данным по разделению дериватизованных гидролизатов bR методом ОФ ВЭЖХ, степени хроматографической чистоты выделенных дейтерий-меченных Dns-Phe-OMe, Dns-Tyr-OMe и Dns-Trp-OMe составили 96, 97 и 98% соответственно.
Исследование степени дейтерироваиности бактериородопсина. Оба подхода показали хорошие результаты по введению дейтериевой метки в молекулу bR. Например, в масс-спектре гидролизата электрофоретически чистого bR, полученного с селективной среды, содержащей L-[2,3,4,5,6-D5]-Phe, L-[3,5-D2]-Tyr и L-2,4,5,6,7-D5]-Trp, после прямой обработки реакционной смеси Dns-Cl и CN2Н2 фиксируются пики, соответствующие молекулярным ионам обогащённых дейтерием Dns-Phe-OMe с М+. при m/z 417 (вместо m/z 412 в контроле), Dns-Tyr-OMe с М+. при m/z 429 (вместо m/z 428) и Dns-Trp-OMe с М+. при m/z 456 {вместо m/z 451).
В случае с униформным мечением bR, метка включалась равномерно по всем положениям углеродного скелета в аминокислотных остатках белка.
ВЫВОДЫ:
1. Подобраны условия для проведения адаптации штаммов R. melhylicum, H.halobium, В. xubiilis и II. amyloliquefaciens к росту на D2О-средах. Селекционно отобраны штаммы, сохранившие высокие ростовые и биосинтетические характеристики на средах с максимальными концентрациями тяжёлой воды.
2. Показана принципиальная возможность использования суммы химических компонентов дейтеро-биомассы факультативных метилотрофных бактерий В. methylicum в качестве источников ростовых субстратов для синтеза дейтерий-меченных БАС.
3. Изучено влияние меченных субстратов – D2O, CD3OD и |3СН3ОН на ростовые и биосинтетические параметры различных штаммов -продуцентов БАС. Показано, что униформные уровни включения дейтерия в молекулы синтезируемых БАС можно получить, используя высокодейтерированные среды (D2О и СНзОН), а в случае с 13С-мечением того же результата можно достигнуть за счёт использования |3СНзОН.